




1 引言
在生物系统中,螺旋运动是许多微生物采用的运动方式[7],因为在低雷诺数条件下生物个体与环境相互作用受粘滞力支配,此时微游泳体的定向移动要遵循扇贝原理 (Scallops Principle)[8],其中大多数微生物游泳体的定向运动由多个周期微小螺旋非互易 (Non-reciprocal) 运动轨迹累积实现。生物游泳体的运动速度一般可分解为旋转速度矢量和平动速度矢量。当个体运动速率恒定时,如果旋转速度矢量与平动速度矢量平行或垂直则游泳轨迹分别呈现直线或圆形;否则其轨迹是螺旋路径,其中心轴平行于身体的净位移方向[9]。由上述运动学分析可知螺旋轨迹是普遍存在的生物运动模式。螺旋运动的非互易特性赋予螺旋运动高效的运动能力[10],有助于生物游泳体在复杂环境中定位环境刺激并快速调整前进方向从而趋利避害,对于实现种群分布、检测营养梯度、躲避捕食者等多种生物功能都至关重要[11⇓~13]。
本文首先归纳了以生物游泳体为灵感来源制备的各种人工游泳体的运动模式和唯象规律,进一步引入各种推动力如外部物理场、界面/表面张力、化学力,分别探讨其驱动原理及各自优点和局限性。最后总结和展望了人工游泳体螺旋运动设计和运动机制等研究面临的困难、挑战以及未来研究方向。
2 生物游泳体的螺旋运动与仿生设计
近些年研究人员对自然界中纤毛/鞭毛驱动生物游泳体运动的特征和功能开展了广泛的研究,它们的运动主要依赖于螺旋状鞭毛或纤毛等微结构的推进[20],类比制备出工艺简单、能够高效推进和精确导航的仿生微型游泳体,在药物的定向运输和环境修护等方面具有特别重要的意义。本节将介绍以鞭毛和纤毛结构为灵感设计的游泳体。
2.1 仿生鞭毛驱动游泳体
许多微小的生物游泳体通过旋转其螺旋状的鞭毛驱动其自身运动,鞭毛推动或拉动微生物细胞的作用形式类似螺旋桨,可实现生物体自身的高效和精准运动[21]。
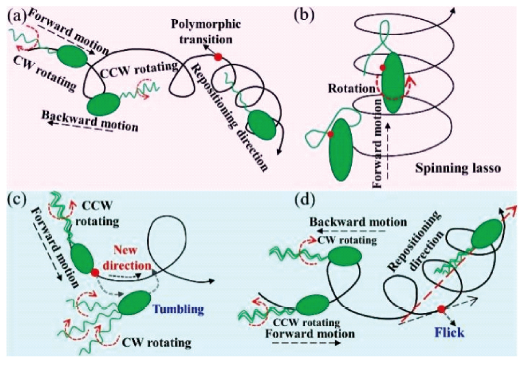
鞭毛的形态和数量可调节生物体的运动功能,具有单鞭毛的细胞通过改变鞭毛的旋转手性控制游动方向。鞭毛逆时针 (Counterclockwise,CCW) 旋转时推动细胞向前移动;鞭毛顺时针 (Clockwise,CW) 旋转时拉动细胞向后移动 (图1 a)[22]。

虽然生物马达的加入为人工游泳体提供了功能更加复杂和强大的运动附件,能够在生物环境中发挥作用的优势。但现有这些附件的稳定性相对较差,持续运动的时间较短,在实际运用过程中的操作较为复杂,合成技术还不够成熟限制了其发展和应用[40]。
2.2 仿生纤毛驱动游泳体
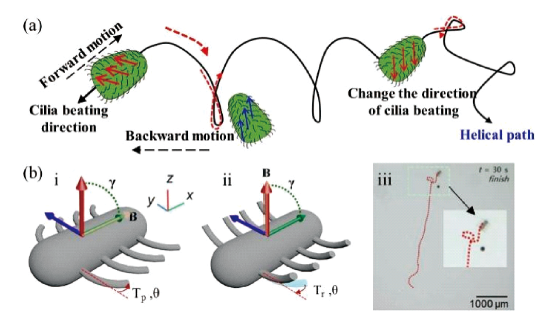
2016年,Choi等[47]提出了一种通过非交互驱动来模拟草履虫纤毛跳动的人工纤毛微机器人 (图3 b),纤毛结构是利用镍沉积的双光子聚合结构,通过逐渐改变磁场方向和强度对功率冲程和恢复冲程进行编程,当被磁场驱动时,纤毛以非可逆方式运动,从而产生净推进力。运动方式由纤毛的形状参数 (例如纤毛长度和纤毛角度) 以及场强和场角度控制,通过这些参数可以精确控制微型机器人的位置和方向。
3 人工游泳体螺旋运动的驱动力和运动控制
大自然创造的高效生物马达可以在复杂的环境中进行运动,真实生物复杂的运动机制可以为人工游泳器和可控装置的开发提供灵感和设计方法。Bibette等[33]提出了实现人工游泳体的可控游泳或运动需要满足的两个条件:首先,能量的供给并转化为系统的机械运动;其次,转化的顺序必须是循环的,而不是时间可逆的。近年来研究者制备出了许多可以模仿生物运动轨迹的人工游泳体,并在人工游泳体的设计和运动行为控制方面提出了多种致动方法。
本文中根据推动人工游泳体产生螺旋运动的能量来源将驱动力分为三类:一类是外部差异物理场力,包括交流电场、磁场、光和超声等;第二类是界面/表面张力;第三类是化学力驱动,例如化学-机械效应等。
3.1 外物理场驱动的螺旋运动
不考虑人工游泳体形状和布朗运动前提下,游泳体在恒定物理场中一般只能进行简单的向前或向后运动,不会呈现复杂运动轨迹。但当环境物理场存在差异时,游泳体会根据物理场的强度或者方向调整自身的运动参数 (速度和方向等) 以适应环境的变化,因此运动行为会变得丰富。
3.1.1 电场驱动游泳体
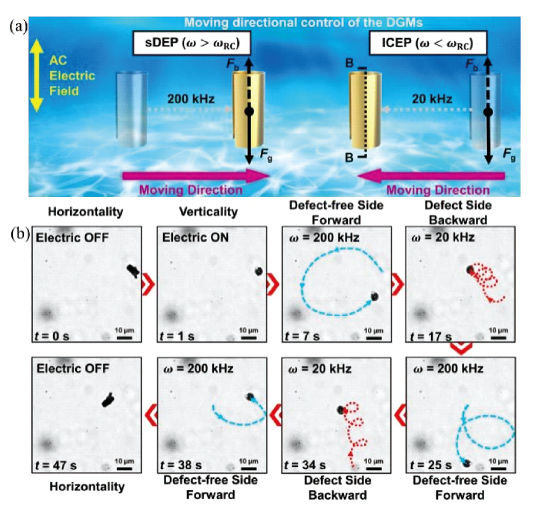
环境电场作为一种外部能源,不仅可以通过多种机制推动人工游泳体产生运动,还可以实现对游泳体位置的控制。利用交流电驱动Janus粒子的运动通常有两种推进机制,即自介电电泳 (sDEP) 和感应电荷电泳 (ICEP)[52]。它们的基本原理分别是通过构造局部电场梯度实现非对称静电力或产生非对称流体在电磁铁周围局部流动。Miloh等[53]提出sDEP推进一般发生在高频交流电场环境,即因Janus粒子两个半球的非对称介电常数引起局部化电梯度倾向于与诱导偶极矩相互作用,产生净静电力驱动Janus粒子向前移动。而ICEP是由感应电荷电渗透 (ICEO) 流在低频交流电场环境[54]下诱导的。相比于sDEP,ICEP可以更加精准控制Janus粒子的运动路线,促使粒子沿着设定的路线运动。
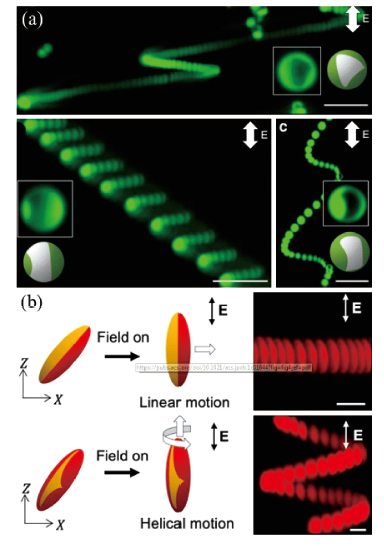
Bharti等[56]近期研究工作发现,该类球型粒子螺旋运动轨迹的几何形状可以由贴片和粒子的几何形状决定。椭球体 (图4 b) 上金属覆盖范围的几何结构与活性粒子进行线性、圆形和螺旋运动等行为之间存在明确的控制规律。结果表明椭球型粒子螺旋运动轨迹的螺距和直径与粒子纵向和横向轴上不对称的金属贴片的特征相关:随着贴片纵向对称性的增加,螺旋螺距普遍减小;随着横向对称性的增加,螺旋直径减小。
3.1.2 磁场驱动游泳体
磁场由于其非侵入性、高穿透性和强可控性,被广泛用作人工游泳体的驱动力,通过控制场强大小和场方向等条件即可控制游泳体运动的行为[57]。在生物游泳体仿生设计中,我们介绍了多种具有螺旋或柔性结构的磁场驱动的仿生游泳体,它们可以随时调整尾巴或头部,以响应磁场的变化,从而向前推进。此外,在微机器人领域,研究者们也制备出了多种磁驱动的螺旋运动机器人。
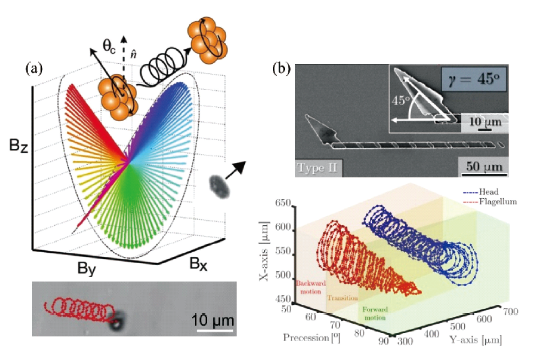
Neeves等[58]探究了一种平面旋转磁场驱动胶体微轮产生旋转耦合的运动机制。通过改变所施加的磁场相位角,可快速、简单地控制微轮的运动方向,使运动控制即时且精确 (图6 a)。实验结果表明通过旋转磁场控制微轮产生的螺旋运动,其靶向性和破除障碍功能均强于直线运动。由于微轮的组装和平移都是通过微弱的、毫微特量级的外部磁场来操纵的,这种磁场不会在组织中衰减,可用于远离身体外表面的阻塞血管的药物输送。
3.1.3 光驱动游泳体
Wang等[63]开发了一种具有光敏性的AgCl-Janus游泳体,在强光照射下,AgCl微粒附近产生离子扩散电泳推动Janus粒子向远离反应 (AgCl) 侧运动,光可诱导Janus粒子以螺旋方式远离重力轴进行运动 (图7 a)。但此类AgCl微马达,具有寿命短、响应时间长等缺点,随着时间的推移其活性化学物质 (AgCl) 持续消耗,因此不可重复使用或可持续使用。
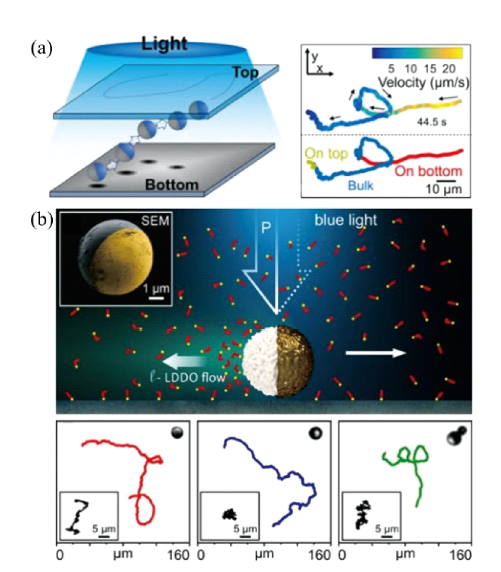
图7 光驱动游泳体的运动行为:(a) 光诱导离子扩散电泳驱动AgCl-Janus 粒子的运动[63];(b) 光诱导扩散渗透流驱动SiO2 颗粒的运动[64]Fig.7 Light-driven kinematic behavior of swimmers: (a) Light-induced ion diffusion electrophoresis driving the motion of AgCl-Janus particles[63]; (b) Light-induced diffusion permeation flow driving the motion of SiO2 particles[64] |
Santer等[64]提出了一种光诱导多孔二氧化硅颗粒自推进运动的机制。在蓝光条件下,当多孔二氧化硅颗粒浸入含有光敏表面活性剂的水溶液,颗粒内会发生光异构化反应,孔内形成的亲水顺式异构体将液体从胶体中排出,孔隙周围产生局部光诱导的扩散渗透 (LDDO) 流,推动胶体粒子产生运动 (图7 b)。这种机制可以简单地通过外部光的强度和波长控制运动方式,从而实现多种运动轨迹。
3.1.4 超声驱动游泳体
Bishop等[66]报道了一种通过超声来远程控制手性胶体游泳体运动行为的方法。他们设计并制造了具有扭曲星形结构的胶体粒子 (图8 a),研究了它们在兆赫频率均匀声场平面内的运动行为。实验中观察到游泳体可以在声场的作用下进行稳定的螺旋运动且旋转方向与胶体的手性形状密切相关。实验结果可以通过稳定流动和超声波致动流体动力学模型进行解释。这项研究提出了通过系统形状和超声场矢量联合控制游泳体复杂运动轨迹的方法。
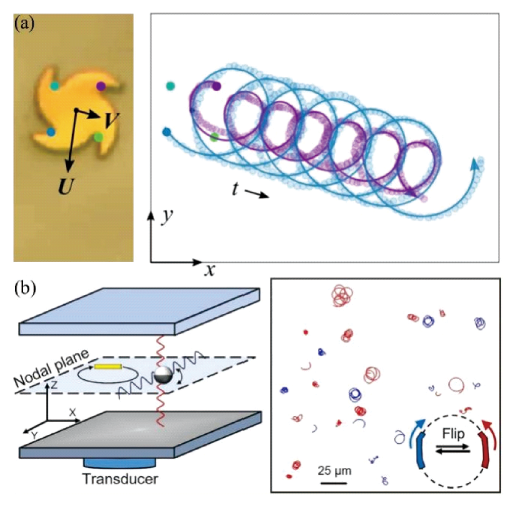
Wang等[67]研究了在兆赫超声波中金属微棒在平面内轨道的运动行为和旋转动力学。实验表明:在共振超声频率附近,这些金属微棒可以沿着紧密的螺旋圈轨迹运行。产生旋转运动行为的可能原因是金属微棒的不均匀性导致金属棒节面上承受的力和扭矩分布不均匀,从而在平面内旋转 (图8 b)。该研究结果为设计和开发具有生物相容性能源和可控运动的游泳体奠定了基础。
以上利用物理场 (电场、超声等) 驱动游泳体进行螺旋运动方面人们已经做出了大量基础性工作,这对于设计复杂环境中航行的微型机器人具有重要的指导意义。但是存在下述问题:人工游泳体的制备过程复杂,制备工艺要求较高,通常需要精确控制特殊的形状和组成,缺乏一种简便、可靠的制造技术,如无外界持续地能量供给则不能进行自主运动,在实际应用中缺乏可控性和远程交互性等等。考虑到实际环境中物理和化学条件的复杂性,对游泳体运动的启动和终止以及实时位置跟踪存在一定困难。
3.2 界面/表面张力驱动的螺旋运动
在外场恒定或无外场的人工系统中推进游泳体运动的另一种方法是仅通过改变游泳体几何形状的不对称性或游泳体与液面的相互作用,由流体流动与局部场梯度的耦合推进游泳体的运动,一般情况下可以通过改变游泳体的物理或化学不对称性来实现[68]。马兰戈尼 (Marangoni) 效应与溶质效应引起的表面物理场的对称性破缺可以推动人工游泳体进行直线、曲线以及螺旋等多种模式的运动,在本小节中重点介绍游泳体的螺旋运动行为。
3.2.1 马兰戈尼效应驱动游泳体
界面作用中通过Marangoni效应产生的剪切力推动游泳体进行螺旋运动最为普遍。由于游泳体形状的非对称性,导致了流体力学意义上的平移-旋转耦合或推进机制中的各向异性[69]。
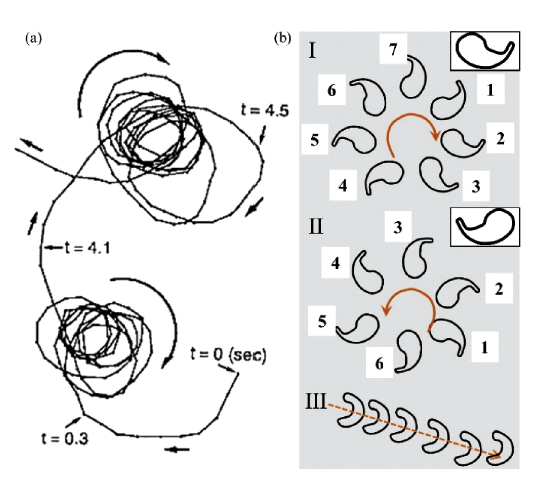
Maass等[71]的工作表明,在不存在手性要素的系统中,人工游泳体的运动轨迹可以简单地通过改变其所处环境的温度进行控制。每个液滴内的向列相序和表面流相互作用可以产生旋转力矩,导致液滴进行二维转圈运动。将液晶液滴加热到35 ℃时,液滴内部的分子变得无序导致旋转力矩消失,液滴恢复到游动状态。如图10 所示,这种运动模式的转变可以系统地和可逆地随温度调节。
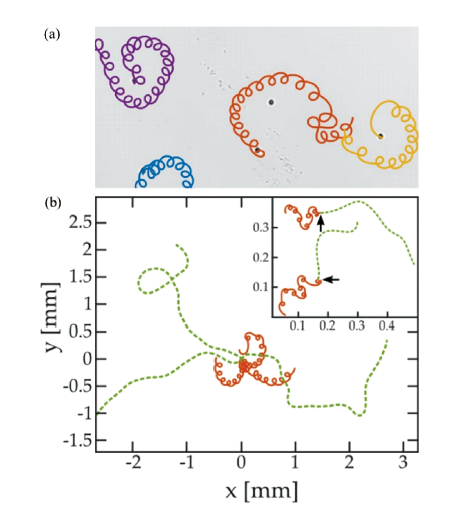
图10 温度控制人工游泳体的运动轨迹转变,液晶滴(向列)在34 ℃的轨迹以实线示出,液晶滴(各向异性)在37 ℃的轨迹以虚线示出[71]Fig.10 Temperature-controlled shifts in the trajectory of artificial swimmers. The trajectory of the liquid crystal droplet (nematic) at a temperature of 34 ℃ is shown as a solid line;the trajectory of the liquid crystal droplet (isotropic) at a temperature of 37 ℃ is shown as a dashed line[71] |
$ \begin{array}{c}\\ \dfrac{\text{d}\nu_i}{\text{d}t}=\gamma p_i-\nu_j\nu_j\nu_i+a_1Q_{ij}\nu_j+\\ \\ a_2\epsilon_{ijk}\omega_j\nu_k+\mu^{iso}+\mu Q_{ij}\omega_j\end{array}$
$ \begin{matrix}\mathrm\\ \dfrac{\mathrm{d}\omega_i}{\mathrm{d}t}=\zeta\omega_i-\omega_i\omega_j\omega_i+b_1Q_j\omega_j+\\ \\ b_2\epsilon_{\text{vjk}}Q_j\nu_l\nu_k+\nu^{is\alpha}\nu_i+\nu Q_j\nu_j\end{matrix}$
$ \dfrac{\mathrm{d}Q_{ij}}{\mathrm{d}t}=\epsilon_{\mathrm{kjl}}Q_{ik}\omega_{l}-\epsilon_{_{ikl}}\omega_{i}Q_{kj}$
通过分析各手性运动之间的分岔行为,发现了速度-角速度手性耦合项、CLC液滴结构各向异性以及模型方程中非线性项对CLC液滴螺旋运动行为起着至关重要的作用,该模型也适用于描述其他类型手性自推进。
3.2.2 界面溶质效应驱动游泳体
3.3 化学力驱动的螺旋运动
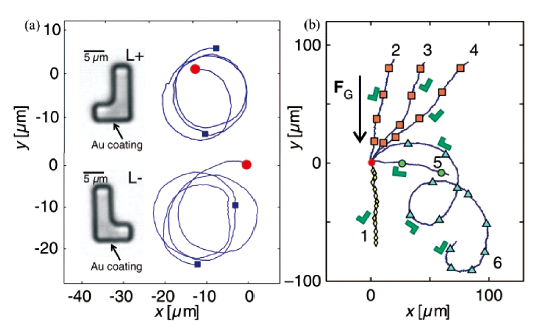
2004年,Crespi等[81]提出通过化学反应驱动且能够自主运动的人工合成游泳体,他们发现了双金属纳米棒在过氧化氢溶液中可以自主移动。该工作促进了通过化学力驱动人工游泳体成为研究热点。
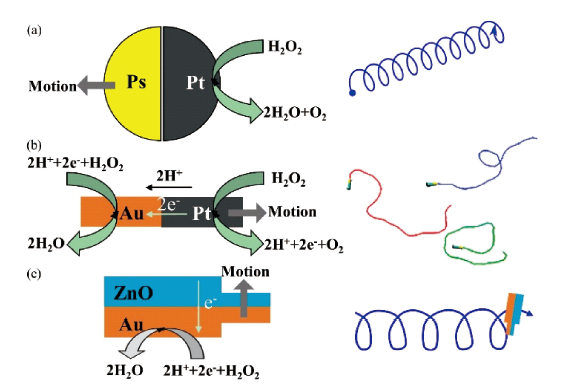
2017年,Ebbens等[85]研究了催化剂表面分布不对称的球形Janus粒子在三维空间 (3D) 中的运动轨迹,Janus粒子一半球面覆盖铂金属催化剂材料 (图13 a),Pt催化过氧化氢水溶液分解为氧和水产生气泡,气泡的喷射将动量传递给游泳体推动Janus粒子运动。由于铂涂层的厚度和胶体的大小的不同使Janus粒子运动时产生了方向偏差,运动轨迹为螺旋状。
自电泳驱动的Janus粒子运动是由于粒子周围电化学反应的不对称性,使得带电粒子在自身产生的电场中移动。Mallouk等[86]开发的Au-Pt双段自电泳纳米棒,可以以大约10 μm/s的速度向Pt端产生定向运动行为 (图13 b)。
Zhang等[87]合成了可进行多种运动模式的不对称ZnO/Au Janus纳米棒 (图13 c)。随着H2O2浓度或紫外光强度的增加,ZnO/Au棒的运动轨迹由弹道运动过渡到持续的旋转运动。通过调节入射光的角度,ZnO/Au棒的运动轨迹可在直线平移、螺旋运动和圆周运动之间可逆转变。
基于Janus粒子结构不对称性引起的力学不稳定性可以设计触发活性旋转,且不需要内置的手性形状或手性化学活性。但是Janus粒子运动存在下述局限性:使用催化分解过氧化氢来实现推进通常会导致不稳定的运动;粒子的旋转自由度缺乏可控制性[88];反应进行中燃料的逐渐消耗可导致游泳体运动能力的减弱。如何实现Janus粒子的可控运动以及持续能量供给将是未来研究的重点。
近期的研究工作表明游泳体形状的非对称不是产生旋转运动的必要条件,即使在没有上述游泳体形状不对称性的情况下,游泳型液滴在非平衡状态下也会自发进行螺旋运动。当液滴能够从内部产生化学分子时,化学反应和运动之间的耦合会自发地破坏对称性,导致液滴的定向运动。表面张力在该类体系中起着至关重要的作用;它也可以用作化学-机械传感器,化学分子的不均匀浓度可转变为液滴表面的机械力学差异从而实现检测过程[89]。
BZ反应液滴内部化学振荡反应进行时其催化剂铁的氧化态比还原态对应的溶液系统具有更高的界面张力。随着液滴内化学振荡反应持续进行,液滴表面张力将会周期性变化。当液滴内部存在周期性化学波传播时则可诱导流体定向流动,从而驱动BZ液滴的运动。Sumino等[90]研究了螺旋波驱动BZ液滴的运动。当BZ液滴内部自发产生螺旋波时,界面张力梯度受螺旋波驱动而旋转,旋转对流根据梯度产生并诱导旋转运动 (如图14 )。实验中观测到BZ液滴的运动形式是短程、随机且不可控的,这是由于目前还没有建立成熟的实验技术可以在BZ液滴中激发和控制能够长时间定向且稳定传播的化学波。
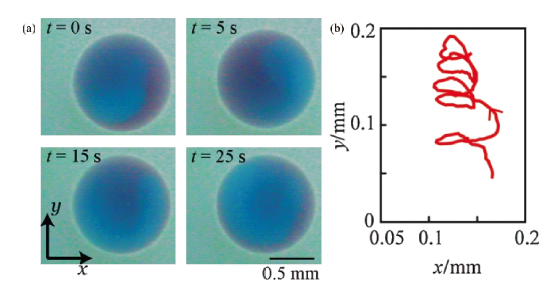
Gao等[102]在2021年提出由一种活性自振荡BZ凝胶驱动的螺旋运动模式,其可以在外界环境均匀且材料自身和边界均对称的情况下,仅通过内部化学波信号的不对称性驱动活性凝胶产生螺旋运动轨迹。研究发现由化学波驱动的BZ凝胶在一维玻璃管中进行的趋光和避光运动[98]与封闭在一维玻璃管中的Euglena gracilis具有类似的向前和向后运动模式[103]。在二维溶液环境中,矩形BZ自振荡凝胶中螺旋波/脉冲波在x-y二维空间上传播,驱动力的方向会偏离波前法向方向。凝胶依次沿着有相位差的法向运动矢量和切向运动矢量交替向前和向后进行逆波运动,形成螺旋运动轨迹 (图15 )。与BZ液滴的运动 (界面张力驱动) 相比,矩形BZ凝胶的螺旋运动可以在没有外部相互作用的情况下发生,即它是由内部驱动的。BZ凝胶的这种运动模式、方向和运动速度可以通过外部控制参数 (如光强、温度和BZ溶液的底物浓度) 进行控制。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
这项工作表明,人工游泳体的螺旋运动可以仅由内部传播的信号驱动,不需要内置的手性形状或手性化学活性,也不需要任何外部非对称环境。在控制外界的条件下,如BZ凝胶的形状、化学波在BZ凝胶上的位置等因素,螺旋运动的范围也可控制。但这类由内信号驱动的软机器人的运动速度与生物游泳体相比相差甚远[104],如何提高BZ凝胶的运动速度、运动稳定性以及化学-机械能持续转化性能是未来发展的关键问题。
综上所述,利用化学力驱动游泳体的运动方法丰富了人工游泳体的“燃料”选择。化学反应网络内驱动性质赋予该类系统以定向性、稳定性和自适应性等仿生功能,并在仿生多模式智能运动方面拥有特色和重大发展潜力。目前广泛使用的燃料分子如H2O2、BZ溶液等虽然能够很好地为运动体系提供能量,但不能用于生物体内。因此发展生物相容性好的燃料分子是人工游泳体未来研究发展的重要方向。
4 结论与展望
生物游泳体优异的运动性和功能性激励着科学家们不断模仿和创造具有生物功能的人工游泳体。本综述主要讨论了进行螺旋运动的生物和人工游泳体的运动分类和原理。阐明和概述了利用外部差异物理场、界面/表面张力和化学力推动人工游泳体螺旋运动的方法和运动机制。
通过对人工游泳体螺旋运动行为前期研究的总结,我们提出了关于此领域未来的发展方向如下:
(1) 目前研究者从机械、材料科学、化学、物理、生物学、控制科学等多个学科理论研究出发对人工游泳产生螺旋运动的原因进行分析解释。急需将不同学科阐述的设计方法和运动机理总结出一个普适的螺旋运动模型并进行理论分析。
(2) 微生物可以在黏性复杂的流体中进行多模式螺旋运动行为并执行复杂功能,而目前人工游泳体研究工作主要聚焦在运动模式和运动机制的研究。仿生功能的开发是人工螺旋运动游泳体发展的重要方向之一,如何提高螺旋游泳体的定向性、自主性、物质输运与释放以及群体能力[60]等问题需要进一步探究。
(3) 现阶段人工游泳体缺乏对外部环境的自适应性,未来在人工系统中如何通过材料科学和化学方法开发可感知外部传感器信号并将信号传输到控制系统以形成闭环反馈来驱动执行器并实现相应的运动的智能游泳体是一个重要研究方向。化学能驱动的游泳体具有独特优势,但如何提高其驱动性能的稳定性和持续性是一个挑战。