




1 引言
电活性菌在新陈代谢过程中将产生的电子传递至胞外电子受体或者接收胞外电子的过程称为胞外电子传递(Extracellular electron transfer, EET)。EET的研究涉及微生物、材料、电化学、环境等多学科交叉,在元素的地球化学循环及微生物群落间的电子通讯过程中起关键作用,在生物能-电能转换、生物浸出、生物电合成及原位环境污染修复等方面具有广阔的应用前景。
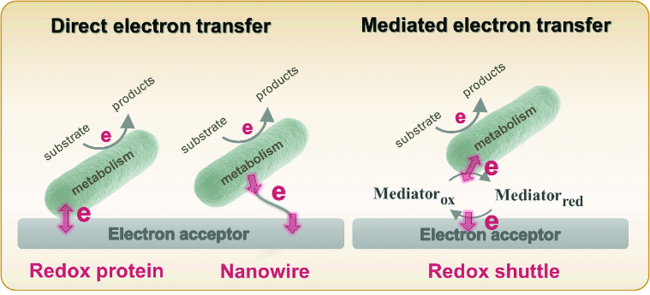
电活性菌的胞外电子传递包括直接电子传递和间接电子传递两种方式(图1)。直接电子传递是指微生物和胞外电子受体间通过直接接触传递电子,有两种形式:(1)近距离电子传递,常指通过微生物外膜/细胞壁上的氧化还原蛋白将电子传递到电子受体,这些蛋白包括细胞色素、黄素蛋白、多血铜蛋白[1,2,3,4];(2)远距离电子传递,在较厚的生物膜中,远离电极的细菌需要借助“纳米导线”或者菌毛将电子传递至电子受体,这一过程需要多步电子跃迁来完成[5, 6]。间接电子传递是指微生物利用新陈代谢产生的初级代谢和次级代谢产物来介导电子传递,这些电子中介体具有氧化还原活性,如Geobacter和Shewanella分泌的核黄素[7, 8]、Pseudomonas aeruginosa分泌的吩嗪[9]和Lactococus lactis分泌的醌类物质[10]等。
早期发现的具有胞外电子传递能力的模式菌Shewanella和Geobacter[11,12,13,14]均属于革兰氏阴性菌,而当前关于革兰氏阳性菌胞外电子传递的研究报道相对较少。由于革兰氏阳性菌细胞壁由90%肽聚糖和10%磷壁酸组成,厚度约为30~80 nm,其结构与革兰氏阴性菌存在明显不同,由此可以推断电子在跨膜传递过程中涉及的蛋白等很可能存在区别。另一方面,已有报道的革兰氏阳性菌可在极端环境中(如高温、高盐、碱性)生存[2, 15, 16],这对拓展胞外电子传递机制的应用具有重要意义。因此,归纳总结革兰氏阳性电活性菌的胞外电子传递机制有助于进一步开展和完善微生物的电子传递研究,拓展微生物的能量和物质转化在环境、能源和材料等领域的应用。
2 革兰氏阳性电活性菌的胞外电子传递
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
2.1 厚壁菌门芽孢杆菌纲
2.1.1 芽孢杆菌目
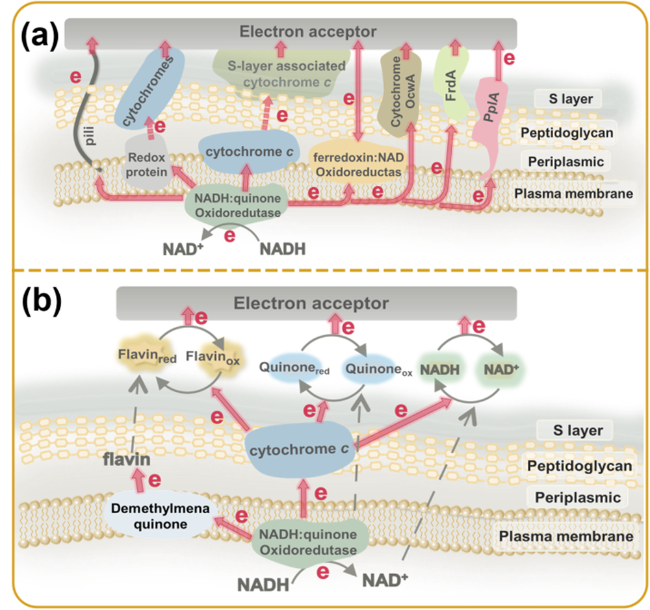
芽孢杆菌纲由芽孢杆菌目和乳酸杆菌目组成,芽孢杆菌目为产芽孢、需氧或兼性厌氧的革兰氏阳性菌,种类繁多,广泛存在于土壤水体中,其可通过膜结合蛋白、固定蛋白或表面蛋白进行直接电子传递。Modestra等[19]发现Bacillus subtilis在0.04 V、0.211 V和0.423 V处有三个氧化还原峰,其对应的膜结合蛋白分别为细胞色素c、bc1和aa3,该菌利用这些膜结合蛋白进行直接电子传递。Listeria monocytogenes为发酵型革兰氏阳性电活性菌,计时电流及循环伏安数据显示该菌具有胞外电子传递能力,研究者发现其利用膜固定脂蛋白PplA上的黄素单核苷酸(FMN)基团将电子转移至胞外,完成直接电子传递[3]。除上述的PplA途径外,Light等[20]发现延胡索酸作为电子受体时,L. monocytogenes的电子传递过程为NAD-醌类物质-延胡索酸,其中醌类物质-延胡索酸是由表面蛋白FrdA介导。通过LC-MS/MS分析发现FrdA黄素化位点Ser48发生变异时,琥珀酸的生成量明显减少,菌体不生长,这说明Ser48的黄素化对FrdA的活性至关重要;同时,发现在L. monocytogenes中重组表达黄素化的胞外尿刊酸酯还原酶,在电子传递过程中的作用与FrdA一致,该研究证明细胞表面黄素化蛋白介导的胞外电子传递是一种重要的直接电子传递途径[20]。
芽孢杆菌目细菌通常也可以利用自身分泌的电子穿梭体进行间接电子传递。Chen等[21]发现B. subtilis在添加愈创木酚和硫酸铜的LB培养基中,培养7 d后具有电化学活性,并且该菌在经过100 ℃高温处理8 h后依然有电化学活性;另外,该菌在酸性条件下(pH=1.5的缓冲液)存放2年后仍具有氧化还原能力,并且能利用乙酸钠产生电子;分泌的烟酰胺腺嘌呤二核苷酸(NAD+)可作为电子中介体在菌体与外界电子通讯中起主要作用;但是极端条件下胞外电子传递在细菌生存中起的作用需要进一步研究。另外,B. megaterium LLD-1[22]和Bacillus sp. WS-XY1[23]均可分泌核黄素并作为电子中介体进行间接电子传递,其中,Bacillus sp. WS-XY1在生长至120 h时分泌0.32 μM核黄素,能够产生约(6.3 ± 2.5)μA的电流。嗜热芽孢杆菌Geobacillus sp. Iso5在低于50 ℃时不具有电活性,在55 ℃该菌产生胞外聚合物后可以检测到电活性,采用高效液相色谱(HPLC)和基质辅助激光解析串联飞行时间质谱(MALDI-TOF)分析其胞外聚合物,结果表明其电活性与核黄素相关[24]。而且,Bacillus cereus DIF1也可以分泌核黄素和黄素单核苷酸介导胞外电子传递应用于微生物燃料电池[25]。由此可见,芽孢杆菌能分泌的电子穿梭体有NAD+、核黄素和黄素单核苷酸,它们在酸性和高温环境中应用具有特定优势。
2.1.2 乳杆菌目
乳杆菌目通常具有嗜酸性,耐酸力较强,目前报道的乳杆菌目电活性菌有Enterococcus faecalis、Enterococcus avium(Gut-S1)、Lactobacillus plantarum和Lactococcus lactis。其中,E.faecalis呼吸链相对简单,Pankratova等[26]发现还原态醌类物质(demethyl menaquinone)在其胞外电子传递过程中起关键作用。添加0.5 mM甲基萘醌前后,E. faecalis产生的电流密度从0.9 μA·cm-2增加至(43.1 ± 1.6) μA·cm-2,另外,在细胞色素bd缺失菌株中检测到的还原态甲基萘醌含量相比野生型菌株的更多,这说明细胞色素bd阻碍甲基萘醌还原。Keogh等[27]发现血红素也影响E. faecalis的胞外电子传递能力,在缺少血红素时,L-乳酸盐脱氢酶通过生物膜固定的铁离子完成并促进胞外电子传递。
Hederstedt等[28]研究E. faecalis OG1RF及其Ndh3和EetA缺陷菌株的电子传递时发现,当铁离子为电子中介体时,其电子传递和铁还原酶活性菌均依赖于膜结合蛋白Ndh3和EetA,然而当锇氧化还原聚合物为中介体时,其电子传递不依赖于这两个蛋白。无论是锇氧化还原聚合物还是铁离子作为中介体其电子传递都依赖于还原态醌类物质。Naradasu等[29]从人体肠道中分离出一株革兰氏阳性电活性菌Gut-S1,当电压设置为0.4 V(vs SHE)时,在灭菌培养基中添加Gut-S1后电流约达到90 nA·cm-2,添加30 mM乙酸电流持续下降,然后添加葡萄糖,电流增至120 nA·cm-2,葡萄糖发酵产物为乳酸和乙酸,说明在该菌中葡萄糖发酵与胞外电子传递结合在一起。更换新鲜培养基后,在短时间内其电流下降20%,然后恢复至原水平,说明可溶性电子中介体对其电流的产生影响有限,其电子传递主要由氧化还原蛋白介导,但蛋白类型未知。因此,在今后的研究中需要考虑微生物代谢对胞外电子传递的影响。
Lactococcus lactis为同型发酵菌,可以产生多种醌类物质来将电子转移至胞外并还原Fe3+、Cu2+等,其中2-氨基-3-乙二酸-1,4萘醌作为电子中介体在电子传递中起主要作用[10]。
2.2 厚壁菌门梭菌纲
梭菌纲和芽孢杆菌纲同属厚壁菌门,其主要区别为芽孢杆菌纲为好氧或者兼性厌氧菌,梭菌纲则为严格厌氧菌,因此研究梭菌纲菌株的电子传递有利于其在厌氧环境中的应用。梭菌纲包括梭菌目和嗜热厌氧菌目,其中梭菌目占主导地位。
2.2.1 梭菌目
梭菌目的直接电子传递通过细胞色素c和铁氧化还原蛋白来实现。从微生物燃料电池中分离出的嗜热菌Thermincola ferriacetica通过在电极表面形成生物膜,将电子直接传递到电极[30]。另一株嗜热菌Thermincola potens通过与细胞壁相连的多血红素细胞色素c进行直接电子传递[2]。Costa等[31]发现Thermincola potens表面非亚铁血红素细胞色素OcwA结构类似于参与氮、硫生物地球循环的多血红素细胞色素c,该蛋白作为呼吸作用中的“瑞士军刀”,无需转录调节或蛋白合成即可迅速选择电子受体以适应环境变化,其可利用的电子受体包括固态物质、可溶性电子中介体(AQDS、核黄素)和含氧阴离子等。
革兰氏阳性电活性菌也包含在地下深处及生态位中占主导地位的硫酸盐还原菌,该类菌具有耐氧、耐干燥的特点。其中,Desulfotomaculum reducens M1能利用膜上细胞色素c552将电子直接传递至胞外受体[32]。Desulfotomaculum ferrireducens GSS09T在以丙酮酸钠作为碳源和电子供体时,通过膜细胞色素c与电子受体直接接触,完成胞外电子传递[33]。Desulfitobacterium dehalogenans以3-氯-4-羟苯基乙酸为电子受体时通过氯氟碳呼吸作用来生长,核黄素蛋白介导醌脱氢酶与跨膜蛋白CprA电子传递,完成该菌的胞外呼吸作用[34]。另外,铁氧化还原蛋白复合体(ferredoxin:NAD oxidoreductase)是厌氧菌中离子驱动的可逆电子转移系统,在厌氧菌(少量古菌)的生物能量学研究中至关重要[35]。Acetobacterium woodii为厌氧产乙酸菌,依赖于跨膜Na+ 梯度产生的膜电势,电子可以在铁氧化还原蛋白与胞外受体之间进行双向传递[35]。Clostridium butyricum EG3为严格厌氧的革兰氏阳性菌,其胞外电子传递可能由细胞色素介导[36],当其以葡萄糖或葡萄糖/Fe(O)OH为底物时,可以产生甲酸盐、乳酸盐、乙酸、H2、CO2、丁酸盐等,可用作其他电活性菌的电子供体。
随着肠道微生物的功能逐渐引起关注,其中有关革兰氏阳性电活性菌的研究也初见报道。例如,从小鼠肠道分离的Clostridium cochlearium以葡萄糖为底物时, 5.2 h产电电流密度可达(0.53 ± 0.02)mA·cm-2,并根据循环伏安曲线的峰电位(0.22 ± 0.05)V(vs Ag/AgCl sat. KCl)判断,可能存在未知电子中介体或者浮游细胞参与电子传递[37]。另外,Khan等[38]发现人体肠道微生物Faecalibacterium prausnitzii具有电子传递能力,虽然其为严格厌氧菌,但是能存活于氧气可以通过上皮细胞扩散进入的肠道黏膜,这是因为它通过间接电子传递利用电子中介体核黄素和硫醇将氧气还原,由此可见,胞外电子传递过程可以使肠道微生物更好地适应生存环境。
2.2.2 嗜热厌氧菌目
2.3 放线菌门和绿弯菌门
放线菌门与厚壁菌门相比,其G+C含量高于厚壁菌门。其中,Corynebacterium glutamicum广泛应用于生产氨基酸(如谷氨酸、赖氨酸),其胞外电子传递途径与氧气浓度相关,在低氧和高氧条件下分别通过细胞色素(Cyt.bc1-aa3)和Cyt.bd传递至O2[41]。由此可见,革兰氏阳性菌的胞外电子传递途径也受环境因素影响。Micrococcus luteus可以在碳电极上催化氧气还原,该过程是由其膜上的氧化还原物质介导[42],但该物质尚未明确。Cellulomonas fimi在0.68 V处能产生氧化电流,但不产生电子中介体,当添加能阻断电子从CoQ传递到细胞色素c的抗霉素A时,0.68 V处的氧化电流消失,因此推断该菌具有通过细胞色素c与受体之间进行直接电子传递的能力[43]。另外,Tian等[25]研究发现在以Rhodococcus ruber DIF2构建的微生物燃料电池中,分泌的核黄素和黄素单核苷酸均可介导间接电子传递。
Ardenticatena maritima 110S为目前报道的仅有的一株属于绿弯菌门的革兰氏阳性电活性菌,其分离于富铁海洋热区,含有绿色的色素,通过表面的细胞色素与铁矿之间进行双向电子传递[44]。由于其还能利用光合作用实现能量和物质转化,因此有望将其用于高温条件下的生物光合成和二氧化碳转化等能源转换体系。
3 应用前景
3.1 污染物的降解和去除
革兰氏阳性电活性菌对环境适应能力强,且多数分离于污染环境,因此,其可用于难降解有机污染物和重金属的去除。
污染物的降解过程往往伴随着电子转移,例如,从被石油污染的土壤中分离出来的Bacillus sp.C8能利用石油烃和硝酸盐分别为电子供体和电子受体,添加表面活性剂和蒽醌-2,6-二磺酸盐(AQDS)后,该菌将石油烃的降解率从67.3%提高至76.7%,AQDS作为电子中介体降低了亚硝酸的积累并提高了石油烃的降解[45]。B. subtilis具有较高的降解酚类物质的能力,将其构建的微生物燃料电池用于2,4-二氯苯酚降解,当阴极液为过硫酸钾时,其最大功率密度为9.5 mW·m-2且2,4-二氯苯酚的降解率可达60%[46],这说明该菌采用直接电子传递的方式将电子传递至电极。因此,在有机污染物降解去除过程中,直接电子传递和间接电子传递途径均可用于提高污染物的降解效率。
环境中的重金属多以离子形态赋存,电活性菌通过胞外电子传递过程可将其还原,革兰氏阳性电活性菌也可应用于重金属去除。例如,分离于土壤微生物燃料电池中的Clostridium sporogenes,能通过细胞色素c介导的直接电子传递过程还原污染土壤中的Cu2+ [47]。Desulfotomaculum reducens能够在丙酸盐发酵培养基中还原U(Ⅵ),其以芽孢产生的H2为电子供体,通过分子量小于3 kD的电子中介体进行间接电子传递[48]。Caldicellulosiruptor saccharolyticus利用中性红为电子中介体,在4 h内能将铬的去除量从11.84 ppm提高至33.66 ppm[49],利用间接电子传递过程加速铬的还原。由此可见,革兰氏阳性电活性菌的直接和间接电子传递均能用于重金属还原。另外,兼性厌氧、耐盐菌Exiguobacterium aestuarii YC211能在pH=5~9、盐度0~15 g L-1条件下还原铬,在外阻为1500 Ω时微生物燃料电池功率密度为(98.3 ± 1.5)mW·m-2;且在铬浓度为2.5~60 mg·L-1范围时与输出电压成线性关系,这将拓展革兰氏阳性电活性菌在微生物传感器领域的应用[15]。
3.2 生物能量转化
电活性菌是微生物电化学系统的重要组成部分,其胞外电子传递能力决定了该系统的性能。B. subtilis在M9培养基中以葡萄糖作为电子供体,运行3个月后最大功率密度为1.05 mW·cm-2,其电能是通过菌体分泌的电子中介体介导产生的,但是其种类还未确定[50]。Samsudeen等[51]从酿酒厂的废水中分离出三株革兰氏阳性菌L. sphaericus SN-1、L. sphaericus SN-2和B. safensis SN-3,它们均可在处理废水的同时产电,其中L. sphaericus SN-2的产电能力最为突出,在pH=8.0条件下最大功率密度可达(123.5 ± 3)mW·m-2。厌氧污泥构建的微生物燃料电池的功率密度为1.82 W·m-3,当添加B. cereus至厌氧污泥后,其功率密度增加至4.83 W·m-3,减少了54%的甲烷产生,该过程是通过电子中介体介导完成的[52]。另外,通过添加100 nM 核黄素作为B. megaterium LLD-1的电子中介体,同时以甘油为底物,微生物燃料电池的功率密度达170 mW·m-2,比未添加核黄素组高4.6倍[22]。嗜碱革兰氏阳性菌Corynebacterium sp. MFC03在葡萄糖为底物的碱性微生物燃料电池中,通过自身分泌的氧化还原物质,使电池的最大功率密度可达7.3 mW·m-2[16]。
3.3 生物制品
通过添加外源性电子中介体,革兰氏阳性电活性菌还可以选择性提高生物制品的生成效率,例如,以10 μM 羟钴胺素作为电子中介体时,Acetobacterium woodii利用果糖为电子供体,能在2.5 d内将470 μM CCl4完全转化,与不添加羟钴胺素相比,转化速率提高了30倍。而且,微生物利用羟钴胺素作为电子中介体还提高了CCl4的矿化程度,增加了14CO2(31%)和其他14C标记的可溶性物质(主要是L-乳酸和乙酸)的含量,并减少了氯仿的生成[55]。
4 结论及展望
综上所述,革兰氏阳性电活性菌的胞外电子传递也可归纳为直接电子传递和间接电子传递两类,但是由于其细胞壁的结构与革兰氏阴性菌不同,电子在跨膜传递的过程中利用的氧化还原活性蛋白等存在差别,尤其是革兰氏阳性菌的肽聚糖和磷壁酸在电子传递中的作用尚不够清晰明确,值得进一步关注。另外,部分革兰氏阳性菌具有在极端条件下生存的优势,基于其物质和能量转换会有更广泛的应用前景。
未来针对革兰氏阳性电活性菌胞外电子传递的研究重点包括以下几个方面:(1)借助组学、分子生物学技术,解析革兰氏阳性电活性菌表面及胞内的细胞色素c的种类和功能,及与胞内代谢途径之间的关联。(2)环境中革兰氏阳性菌和革兰氏阴性菌往往共存,当Shewanella oneidensis MR-1与B. subtilis RH33共生于微生物燃料电池时,S. oneidensis MR-1能利用B. subtilis RH33产生的核黄素增强间接电子传递能力,将最大功率密度提高至277.4 mW·m-2[56]。当鼠李糖脂和Pseudomonas sp. CMR12a分泌的1-吩嗪甲酰胺同时存在时,Brevibacillus sp. PTH1利用乙酸钠的产电能力也大大提高,并将Fe3+还原为水铁矿[57]。革兰氏阳性菌与革兰氏阴性菌之间的电子转移相互作用需要进一步研究。(3)大部分的致病菌或肠道微生物属于厚壁菌门革兰氏阳性菌,存在进行胞外电子传递的同源基因[3],通过调控其胞外电子传递或活性,可以减少其对人体的危害。(4)深入了解革兰氏阳性电活性菌的胞外电子传递机制,可以促进其对与生命科学相关的其他学科的理解,如以生物通讯和电子元件为基础的“大脑-机械”人工智能方面的应用[58]。