
文章编号: 1609122939987-537828232
文献标识码: A
FAHFAs:生物功能、分析及合成
收稿日期:2020-07-20
修回日期:2020-11-18
网络出版日期:2020-12-28
基金资助
国家自然科学基金项目(21635006)
国家自然科学基金项目(31670373)
国家自然科学基金项目(21721005)
国家自然科学基金项目(21904099)
中国博士后科学基金项目(2018M642893)
FAHFAs: Biological Functions, Analysis and Synthesis
Received:20 Jul. 2020
Revised:18 Nov. 2020
Online:28 Dec. 2020
Fund
National Natural Science Foundation of China(21635006)
National Natural Science Foundation of China(31670373)
National Natural Science Foundation of China(21721005)
National Natural Science Foundation of China(21904099)
China Postdoctoral Science Foundation(2018M642893)
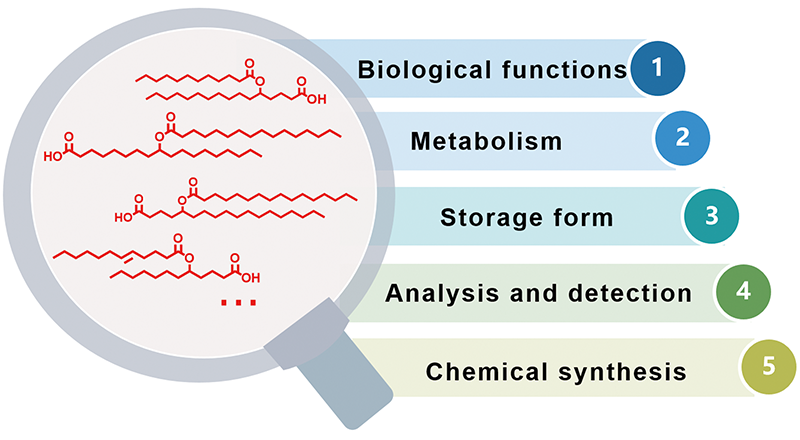
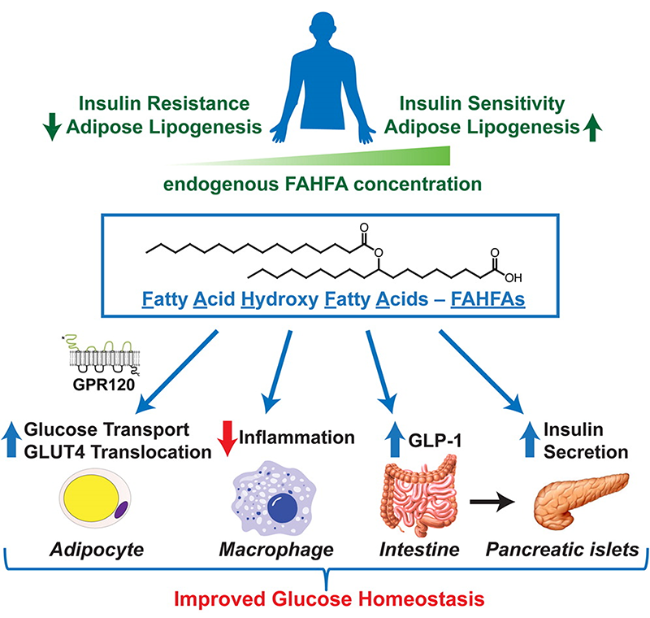
羟基脂肪酸支链脂肪酸酯(branched fatty acid esters of hydroxy fatty acids,FAHFAs)是一类新发现的生物活性脂质分子。FAHFAs在哺乳动物体内具有重要的生理功能,如改善葡萄糖耐量、增强胰岛素敏感性、维持血糖稳态和抗炎等。目前,FAHFA相关研究已成为脂质领域研究的一个新方向,受到科学家们的关注。本文总结了自FAHFAs发现至今的主要研究成果,从FAHFA的生理功能、代谢、生物体内的储存形式、分析检测和化学合成等五个方面进行介绍,以期为未来FAHFAs领域的研究提供一些借鉴。
关键词: 羟基脂肪酸支链脂肪酸酯 ; 生物功能 ; 分析检测 ; 化学合成
中图分类号: O623.61 ()
朱泉霏 , 郝俊迪 , 严靖雯 , 王雨 , 冯钰锜 . FAHFAs:生物功能、分析及合成[J]. 化学进展, 2021 , 33(7) : 1115 -1125 . DOI: 10.7536/PC200741
Quanfei Zhu , Jundi Hao , Jingwen Yan , Yu Wang , Yuqi Feng . FAHFAs: Biological Functions, Analysis and Synthesis[J]. Progress in Chemistry, 2021 , 33(7) : 1115 -1125 . DOI: 10.7536/PC200741
Branched fatty acid esters of hydroxy fatty acids (FAHFAs) are a new class of functional lipids. It has been reported that they play important roles in mammals, such as improving glucose tolerance, enhancing insulin sensitivity, maintaining blood glucose homeostasis, and anti-inflammatory. Since its discovery in 2014, FAHFA has been attracting more and more attention of scientists, and has become a new branch of lipid research. This review summarizes the research advances of FAHFA since its discovery, covering the physiological function, metabolism, storage in organism, analysis and chemical synthesis of FAHFA.
1 Introduction
2 Advances since the discovery of FAHFAs
2.1 FAHFAs biological functions
2.2 FAHFAs metabolism
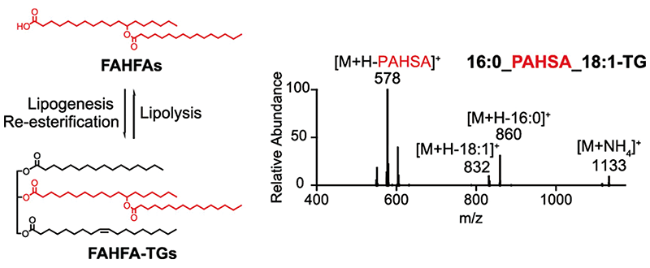
2.3 Storage forms of FAHFAs in vivo
2.4 FAHFAs analysis
2.5 FAHFAs synthesis
3 Conclusion and outlook
Key words: FAHFAs ; biological function ; analysis and detection ; chemical synthesis
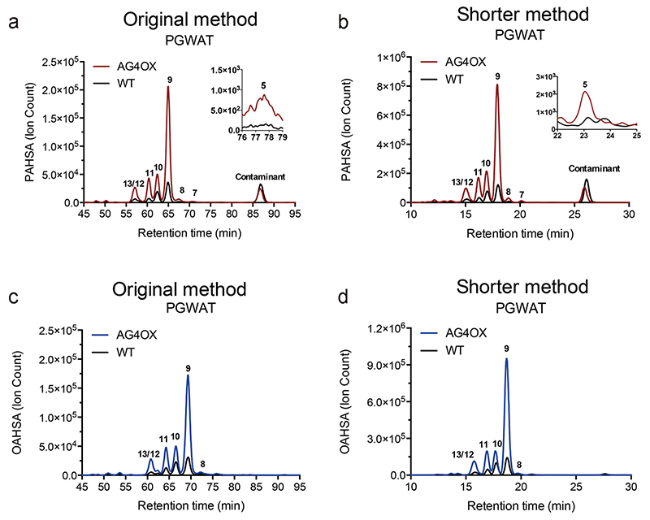
图4 使用原始方法和快速方法分析野生型和AG4OX 型小鼠的腹膜白色脂肪中的OAHSAs和PAHSAs。原始方法(a)和快速方法(b)检测PAHSAs的提取离子流图;原始方法(c)和快速方法(d)检测的OAHSAs的提取离子流图[24]Fig. 4 Analysis of OAHSAs and PAHSAs in PG WAT of WT and AG4OX mice with the original and shorter method[24]. Extracted ion chromatograms comparing perigonadal white adipose tissue (PG WAT) PAHSAs in AG4OX and WT mice using the original (a) and shorter method (b). Extracted ion chromatograms analyzing OAHSAs in PG WAT WT and AG4OX mice in the original (c) and shorter method (d). Copyright 2018, American Chemical Society |
表1 已报道的FAHFAs家族Table 1 Reported FAHFA families |
| No. | FAHFA family | Structure a | Formula | No. | FAHFA family | Structure a | Formula |
|---|---|---|---|---|---|---|---|
| 1 | POHPO | 16:1-O-16:1 | C32H58O4 | 39 | MAHAA | 14:0-O-20:0 | C34H66O4 |
| 2 | OAHPO | 18:1-O-16:1 | C34H62O4 | 40 | PAHAA | 16:0-O-20:0 | C36H70O4 |
| 3 | PAHPO | 16:0-O-16:1 | C32H60O4 | 41 | SAHAA | 18:0-O-20:0 | C38H74O4 |
| 4 | SAHPO | 18:0-O-16:1 | C34H64O4 | 42 | OAHAA | 18:1-O-20:0 | C38H72O4 |
| 5 | LAHPO | 18:2-O-16:1 | C34H60O4 | 43 | LAHAA | 18:2-O-20:0 | C38H70O4 |
| 6 | POHOA | 16:1-O-18:1 | C34H62O4 | 44 | AAHAA | 20:0-O-20:0 | C40H78O4 |
| 7 | OAHOA | 18:1-O-18:1 | C36H66O4 | 45 | PAHDDA | 16:0-O-12:0 | C28H54O4 |
| 8 | PAHOA | 16:0-O-18:1 | C34H64O4 | 46 | PDAHDA | 15:0-O-10:0 | C25H48O4 |
| 9 | SAHOA | 18:0-O-18:1 | C36H68O4 | 47 | PAHDA | 16:0-O-10:0 | C26H50O4 |
| 10 | ALAHOA | 18:3-O-18:1 | C36H62O4 | 48 | HDAHDA | 17:0-O-10:0 | C27H52O4 |
| 11 | LAHOA | 18:2-O-18:1 | C36H64O4 | 49 | SAHDA | 18:0-O-10:0 | C28H54O4 |
| 12 | POHPA | 16:1-O-16:0 | C32H60O4 | 50 | PDAHCA | 15:0-O-8:0 | C23H44O4 |
| 13 | OAHPA | 18:1-O-16:0 | C34H64O4 | 51 | PAHCA | 16:0-O-8:0 | C24H46O4 |
| 14 | PAHPA | 16:0-O-16:0 | C32H62O4 | 52 | HDAHCA | 17:0-O-8:0 | C25H48O4 |
| 15 | SAHPA | 18:0-O-16:0 | C34H66O4 | 53 | SAHCA | 18:0-O-8:0 | C26H50O4 |
| 16 | MAHPA | 14:0-O-16:0 | C30H58O4 | 54 | PDAHHA | 15:0-O-6:0 | C21H40O4 |
| 17 | MOHPA | 14:1-O-16:0 | C30H56O4 | 55 | PAHHA | 16:0-O-6:0 | C22H42O4 |
| 18 | PDAHPA | 15:0-O-16:0 | C31H60O4 | 56 | HDAHHA | 17:0-O-6:0 | C23H44O4 |
| 19 | HDAHPA | 17:0-O-16:0 | C33H64O4 | 57 | SAHHA | 18:0-O-6:0 | C24H46O4 |
| 20 | LAHPA | 18:2-O-16:0 | C34H62O4 | 58 | NAHHA | 19:0-O-6:0 | C25H48O4 |
| 21 | ALAHPA | 18:3-O-16:0 | C34H60O4 | 59 | AAHHA | 20:0-O-6:0 | C26H50O4 |
| 22 | AAHPA | 20:0-O-16:0 | C36H70O4 | 60 | MAHMA | 14:0-O-14:0 | C28H54O4 |
| 23 | POHSA | 16:1-O-18:0 | C34H64O4 | 61 | MOHMA | 14:1-O-14:0 | C28H52O4 |
| 24 | OAHSA | 18:1-O-18:0 | C36H68O4 | 62 | PDAHMA | 15:0-O-14:0 | C29H56O4 |
| 25 | PAHSA | 16:0-O-18:0 | C34H66O4 | 63 | PDEAHMA | 15:1-O-14:0 | C29H54O4 |
| 26 | SAHSA | 18:0-O-18:0 | C36H70O4 | 64 | PAHMA | 16:0-O-14:0 | C30H58O4 |
| 27 | MAHSA | 14:0-O-18:0 | C32H62O4 | 65 | POHMA | 16:1-O-14:0 | C30H56O4 |
| 28 | PDAHSA | 15:0-O-18:0 | C33H64O4 | 66 | SAHMA | 18:0-O-14:0 | C32H62O4 |
| 29 | HDAHSA | 17:0-O-18:0 | C35H68O4 | 67 | FAHFA(38:3) | 20:3-O-18:0 | C38H68O4 |
| 30 | LAHSA | 18:2-O-18:0 | C36H66O4 | 68 | FAHFA(38:5) | 20:4-O-18:1 | C38H64O4 |
| 31 | AAHSA | 20:0-O-18:0 | C38H74O4 | 69 | FAHFA(40:5) | 20:3-O-20:2 | C40H68O4 |
| 32 | ALAHSA | 18:3-O-18:0 | C36H64O4 | 70 | FAHFA(40:6) | 22:5-O-18:1 | C40H66O4 |
| 33 | DHAHLA | 22:6-O-18:2 | C40H62O4 | 71 | FAHFA(40:7) | 20:4-O-20:3 | C40H64O4 |
| 34 | POHLA | 16:1-O-18:2 | C34H60O4 | 72 | FAHFA(42:6) | 20:3-O-22:3 | C42H70O4 |
| 35 | PAHLA | 16:0-O-18:2 | C34H62O4 | 73 | FAHFA(42:8) | 20:4-O-22:4 | C42H66O4 |
| 36 | ALAHLA | 18:3-O-18:2 | C36H60O4 | 74 | FAHFA(42:9) | 20:4-O-22:5 | C42H64O4 |
| 37 | LAHLA | 18:2-O-18:2 | C36H62O4 | 75 | FAHFA(44:9) | 22:5-O-22:4 | C44H68O4 |
| 38 | DHAHDHA | 22:6-O-22:6 | C44H62O4 |
a: The chemical structure of FAHFAs characterized by the carbon number and degree of unsaturation of fatty acids and hydroxy fatty acids. e.g.,16:0-O-18:0 means palmitic acid-hydroxy stearic acid (PAHSA) |
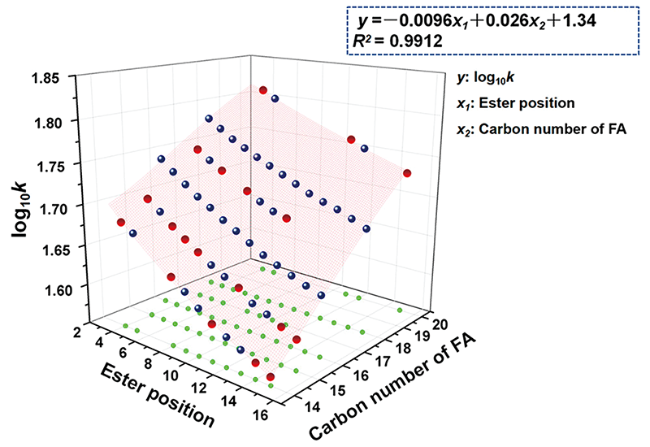
图8 基于FA碳数、酯键位置和log10k的饱和FAHSAs预测模型:其中蓝色圆点代表已确认的化合物,红点代表预测化合物[31]Fig. 8 Prediction model, based on the log10k, ester position, and carbon number of FA, for saturated FAHSAs. Blue dots represent the confirmed compounds; red dots represent the predicted compounds[31]. Copyright 2018, American Chemical Society |
| [1] |
Fahy E, Subramaniam S, Brown H A, Glass C K, Merrill A H, Murphy R C, Raetz C R, Russell D W, Seyama Y, Shaw W. Eur. J. Lipid Sci. Tech., 2005, 107:337.
|
| [2] |
Yore M M, Syed I, Moraes-Vieira P M, Zhang T J, Herman M A, Homan E A, Patel R T, Lee J, Chen S L, Peroni O D, Dhaneshwar A S, Hammarstedt A, Smith U, McGraw T E, Saghatelian A, Kahn B B. Cell, 2014, 159(2):318.
|
| [3] |
Muoio D M, Newgard C B. Nature, 2014, 516(7529):49.
DOI: 10.1038/nature14070 |
| [4] |
Ussar S, Tschöp M H. Cell, 2014, 159(2):238.
|
| [5] |
Lee J, Moraes-Vieira P M, Castoldi A, Aryal P, Yee E U, Vickers C, Parnas O, Donaldson C J, Saghatelian A, Kahn B B. J. Biol. Chem., 2016, 291(42):22207.
|
| [6] |
Kuda O, Brezinova M, Rombaldova M, Slavikova B, Posta M, Beier P, Janovska P, Veleba J, Kopecky J, Kudova E, Pelikanova T, Kopecky J. Diabetes, 2016, 65(11):3516.2.
DOI: 10.2337/db16-er11b |
| [7] |
Kolar M J, Konduri S, Chang T N, Wang H J, McNerlin C, Ohlsson L, Härröd M, Siegel D, Saghatelian A. J. Biol. Chem., 2019, 294(27):10698.
|
| [8] |
Syed I, Lee J, Moraes-Vieira P M, Donaldson C J, Sontheimer A, Aryal P, Wellenstein K, Kolar M J, Nelson A T, Siegel D, Mokrosinski J, Farooqi I S, Zhao J J, Yore M M, Peroni O D, Saghatelian A, Kahn B B. Cell Metab., 2018, 27(2):419.
|
| [9] |
Moraes-Vieira P M, Saghatelian A, Kahn B B. Diabetes, 2016, 65(7):1808.
DOI: 10.2337/db16-0221 PMID: 27288004
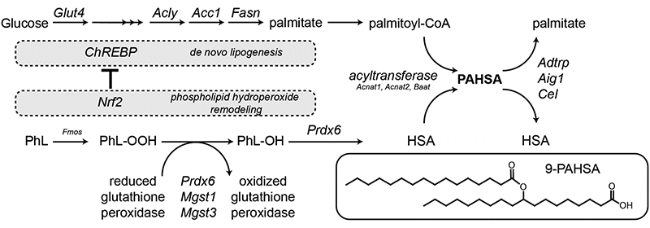
Adipose tissue (AT) regulates systemic insulin sensitivity through multiple mechanisms, and alterations in de novo lipogenesis appear to contribute. Mice overexpressing GLUT4 in adipocytes (AG4OX) have elevated AT lipogenesis and enhanced glucose tolerance despite being obese and having elevated circulating fatty acids. Lipidomic analysis of AT identified a structurally unique class of lipids, branched fatty acid esters of hydroxy-fatty acids (FAHFAs), which were elevated in AT and serum of AG4OX mice. Palmitic acid esters of hydroxy-stearic acids (PAHSAs) are among the most upregulated FAHFA families in AG4OX mice. Eight PAHSA isomers are present in mouse and human tissues. PAHSA levels are reduced in insulin resistant people, and levels correlate highly with insulin sensitivity. PAHSAs have beneficial metabolic effects. Treatment of obese mice with PAHSAs lowers glycemia and improves glucose tolerance while stimulating glucagon-like peptide 1 and insulin secretion. PAHSAs also reduce inflammatory cytokine production from immune cells and ameliorate adipose inflammation in obesity. PAHSA isomer concentrations are altered in physiological and pathophysiological conditions in a tissue- and isomer-specific manner. The mechanisms most likely involve changes in PAHSA biosynthesis, degradation, and secretion. The discovery of PAHSAs reveals the existence of previously unknown endogenous lipids and biochemical pathways involved in metabolism and inflammation, two fundamental physiological processes.© 2016 by the American Diabetes Association. Readers may use this article as long as the work is properly cited, the use is educational and not for profit, and the work is not altered.
|
| [10] |
Syed I, Rubin de Celis M F, Mohan J F, Moraes-Vieira P M, Vijayakumar A, Nelson A T, Siegel D, Saghatelian A, Mathis D, Kahn B B. J. Clin. Investig., 2019, 129(9):3717.
DOI: 10.1172/JCI122445 |
| [11] |
Kahn B B, Saghatelian A, Syed I. EP, EP, US 2019/0151276 A1, 2019.
|
| [12] |
Hamad A R A, Sadasivam M, Rabb H. J. Clin. Investig., 2019, 129(9):3527.
DOI: 10.1172/JCI130313 |
| [13] |
Brezinova M, Kuda O, Hansikova J, Rombaldova M, Balas L, Bardova K, Durand T, Rossmeisl M, Cerna M, Stranak Z, Kopecky J. Biochim. Et Biophys. Acta BBA Mol. Cell Biol. Lipids, 2018, 1863(2):126.
|
| [14] |
Rodríguez J P, Guijas C, Astudillo A M, Rubio J M, Balboa M A, Balsinde J. Cancers, 2019, 11(4):524.
|
| [15] |
Liu T, Tan Z R, Yu J, Peng F, Guo J W, Meng W H, Chen Y, Rao T, Liu Z Q, Peng J B. Expert. Rev. Proteom., 2020, 17(3):233.
|
| [16] |
Zhu Q F, Yan J W, Gao Y, Zhang J W, Yuan B F, Feng Y Q. J. Chromatogr. B, 2017, 1061/1062: 34.
|
| [17] |
Zhu Q F, Yan J W, Ni J, Feng Y Q. BBA-Mol. Cell Biol. L., 2020, 1865: 158639.
|
| [18] |
Parsons W H, Kolar M J, Kamat S S, Iii A B C, Hulce J J, Saez E, Kahn B B, Saghatelian A, Cravatt B F. Nat. Chem. Biol., 2016, 12(5):367.
|
| [19] |
Kolar M J, Kamat S S, Parsons W H, Homan E A, Maher T, Peroni O D, Syed I, Fjeld K, Molven A, Kahn B B, Cravatt B F, Saghatelian A. Biochemistry, 2016, 55(33):4636.
|
| [20] |
Kuda O, Brezinova M, Silhavy J, Landa V, Zidek V, Dodia C, Kreuchwig F, Vrbacky M, Balas L, Durand T, Hübner N, Fisher A B, Kopecky J, Pravenec M. Diabetes, 2018, 67(6):1190.
DOI: 10.2337/db17-1087 |
| [21] |
McLean S, Davies N W, Nichols D S, McLeod B J. Lipids, 2015, 50(6):591.
|
| [22] |
Tan D, Ertunc M E, Konduri S, Zhang J, Pinto A M, Chu Q, Kahn B B, Siegel D, Saghatelian A. J. Am. Chem. Soc., 2019, 141(22):8798.
DOI: 10.1021/jacs.9b00045 |
| [23] |
Zhang T J, Chen S L, Syed I, Ståhlman M, Kolar M J, Homan E A, Chu Q, Smith U, Borén J, Kahn B B, Saghatelian A. Nat. Protoc., 2016, 11(4):747.
|
| [24] |
Kolar M J, Nelson A T, Chang T N, Ertunc M E, Christy M P, Ohlsson L, Härröd M, Kahn B B, Siegel D, Saghatelian A. Anal. Chem., 2018, 90(8):5358.
|
| [25] |
López-Bascón M A, Calderón-Santiago M, Priego-Capote F. Anal. Chimica Acta, 2016, 943:82.
|
| [26] |
Qi B L, Liu P, Wang Q Y, Cai W J, Yuan B F, Feng Y Q. Trac Trends Anal. Chem., 2014, 59:121.
|
| [27] |
Zhang T Y, Li S, Zhu Q F, Wang Q, Hussain D, Feng Y Q. Trac Trends Anal. Chem., 2019, 119:115608.
|
| [28] |
Hu C F, Wang M, Duan Q, Han X L. Anal. Chimica Acta, 2020, 1105: 105.
|
| [29] |
Liberati-Čizmek A M, Biluš M, Brkić A L, Barić I C, Bakula M, Hozić A, Cindrić M. Plant Foods Hum. Nutr., 2019, 74(2):235.
|
| [30] |
Ma Y, Kind T, Vaniya A, Gennity I, Fahrmann J F, Fiehn O. J. Cheminformatics, 2015, 7(1):1.
|
| [31] |
Zhu Q F, Yan J W, Zhang T Y, Xiao H M, Feng Y Q. Anal. Chem., 2018, 90(16):10056.
|
| [32] |
Hu T, Lin M, Zhang D, Li M L, Zhang J L. Anal. Bioanal. Chem., 2018, 410(28):7415.
|
| [33] |
Pham T, Vidal N, Manful C, Fillier T, Pumphrey R, Doody K, Thomas R. Molecules, 2019, 24(2):232.
|
| [34] |
Marshall D L, Saville J T, Maccarone A T, Ailuri R, Kelso M J, Mitchell T W, Blanksby S J. Rapid Commun. Mass Spectrom., 2016, 30(21):2351.
DOI: 10.1002/rcm.7715 |
| [35] |
Ding J, Kind T, Zhu Q F, Wang Y, Yan J W, Fiehn O, Feng Y Q. Anal. Chem., 2020, 92(8):5960.
DOI: 10.1021/acs.analchem.0c00172 PMID: 32202765
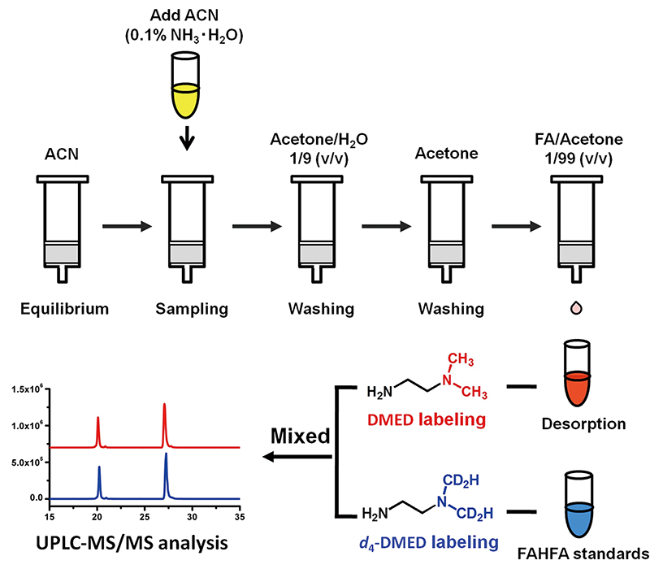
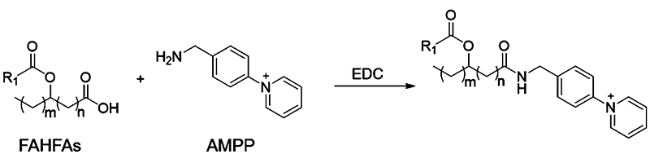
Fatty acid esters of hydroxy fatty acids (FAHFAs) are a family of recently discovered lipids with important physiological functions in mammals and plants. However, low detection sensitivity in negative ionization mode mass spectrometry makes low-abundance FAHFA challenging to analyze. A 2-dimethylaminoethylamine (DMED) based chemical derivatization strategy was recently reported to improve the MS sensitivity of FAHFAs by labeling FAHFAs with a positively ionizable tertiary amine group. To facilitate reliable, high-throughput, and automatic annotation of these compounds, a DMED-FAHFA in silico library containing 4290 high-resolution tandem mass spectra covering 264 different FAHFA classes was developed. The construction of the library was based on the heuristic information from MS/MS fragmentation patterns of DMED-FAHFA authentic standards, and then, the patterns were applied to computer-generated DMED-FAHFAs. The developed DMED-FAHFA in silico library was demonstrated to be compatible with library search software NIST MS Search and the LC-MS/MS data processing tool MS-DIAL to guarantee high-throughput and automatic annotations. Applying the in silico library in samples for profiling FAHFAs by high-resolution LC-MS/MS enabled the annotation of 19 DMED-FAHFAs from 16 families, including 3 novel compounds. Using the in silico library largely decreased the false-positive annotation rate in comparison to low-resolution LC-MS/MS. The developed library, MS/MS spectra, and development templates are freely available for commercial and noncommercial use at https://zenodo.org/record/3606905.
|
| [36] |
Nelson A T. Doctoral Dissertation of University of Texas at Austin, 2018.
|
| [37] |
Balas L, Bertrand-Michel J, Viars F, Faugere J, Lefort C, Caspar-Bauguil S, Langin D, Durand T. Org. Biomol. Chem., 2016, 14(38):9012.
DOI: 10.1039/C6OB01597B |
| [38] |
Nelson A T, Kolar M J, Chu Q, Syed I, Kahn B B, Saghatelian A, Siegel D. J. Am. Chem. Soc., 2017, 139(13):4943.
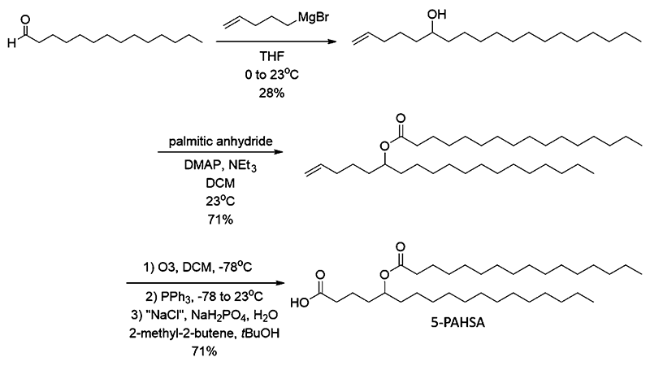
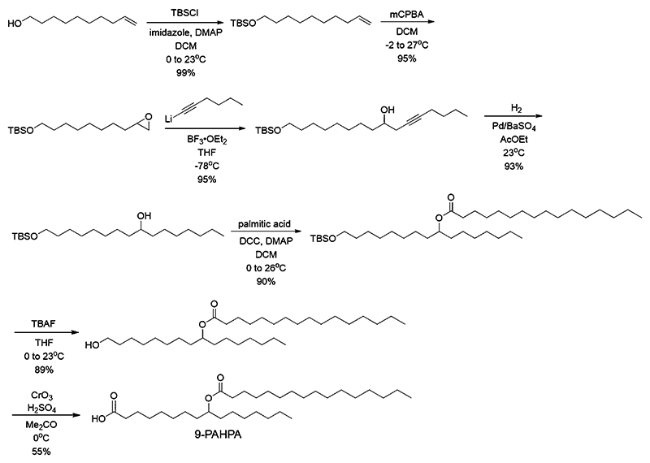
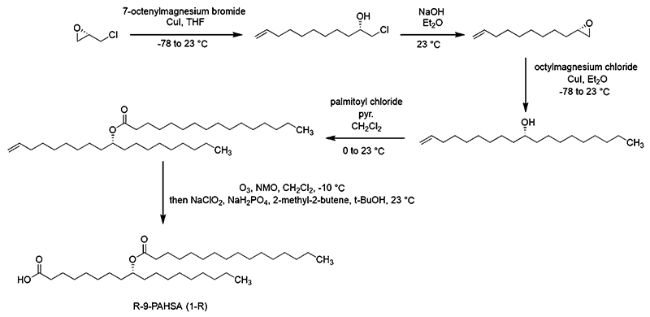
DOI: 10.1021/jacs.7b01269 |
/
| 〈 |
|
〉 |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}