
文章编号: 1608628823751-399889850
文献标识码: A
小分子荧光探针在绿色农药开发中的应用
收稿日期:2020-06-22
修回日期:2020-08-04
网络出版日期:2020-12-22
基金资助
国家重点研发计划项目(2017YFD0201403)
Application of Small-Molecule Fluorescent Probes in the Development of Green Pesticides
Received:22 Jun. 2020
Revised:4 Aug. 2020
Online:22 Dec. 2020
Fund
National Key Research and Development Project(2017YFD0201403)
小分子荧光探针以其灵敏度高、特异性强、稳定性好、操作便捷和成本低等特点在生命科学、医药化学和环境科学等领域得到了广泛的应用。在农药化学领域,小分子荧光探针常被用作农药残留及重金属污染的检测手段。近年来随着全球开发绿色农药战略需求的不断增强,作为靶向型药物设计和高通量筛选的重要分子工具,荧光探针在绿色农药新产品研发领域的应用不断普及和深化。本文从探针分子的化学设计、靶点识别及药物筛选的角度出发,围绕不同类型的绿色农药重要生物靶点,综述了小分子荧光探针在绿色农药开发领域的研究现状,并对其未来的发展趋势和应用前景进行了展望。

侯晓涵 , 刘胜男 , 高清志 . 小分子荧光探针在绿色农药开发中的应用[J]. 化学进展, 2021 , 33(6) : 1035 -1043 . DOI: 10.7536/PC200659
Xiaohan Hou , Shengnan Liu , Qingzhi Gao . Application of Small-Molecule Fluorescent Probes in the Development of Green Pesticides[J]. Progress in Chemistry, 2021 , 33(6) : 1035 -1043 . DOI: 10.7536/PC200659
Small-molecule fluorescent probes are widely applied in the fields of life science, medicinal chemistry and environmental science, due to their characteristics of high sensitivity and specificity, good stability and economic applicability. In pesticide chemistry, small-molecule fluorescent probes are frequently utilized in the detection of pesticide residues and heavy metal pollutions. With global strategic needs and rapid technological progress in green pesticide development, fluorescent probes are urgently desired as important molecular tools for design, screening and development of environmentally benign agrichemicals. This article aims to review the key updates of small-molecule fluorescent bioprobes in green pesticide R&D by covering their chemical design, molecular targeting, and screening mechanisms against different green pesticide biotargets, and to provide the current status on their research and application as well as future perspectives.
1 Introduction
2 Ryanodine receptor-targeted small-molecule fluorescent probes
2.1 Anthranilic diamide-based fluorescent probes
2.2 Phthalic diamide-based fluorescent probes
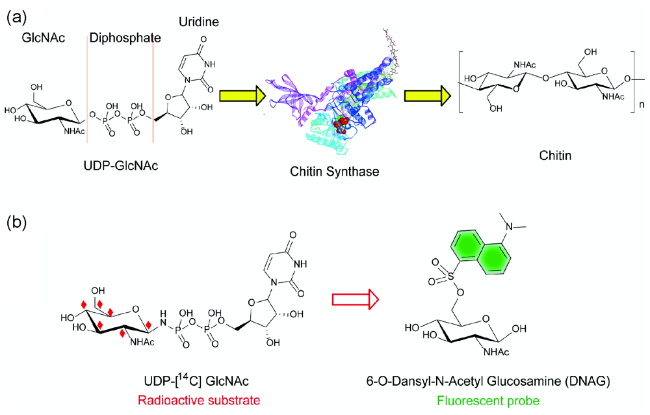
3 Chitin-targeted small molecule fluorescent probes
3.1 Chitin synthase-targeted fluorescent probes
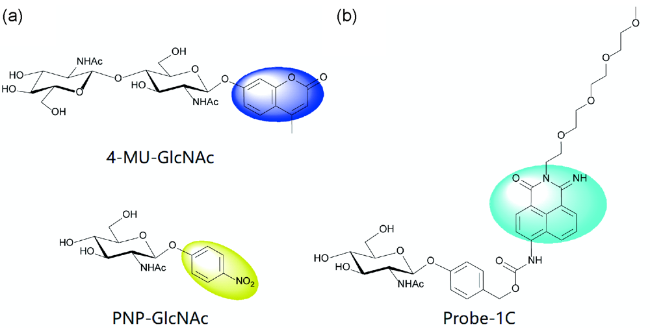
3.2 Chitin related enzyme-targeted fluorescent probes
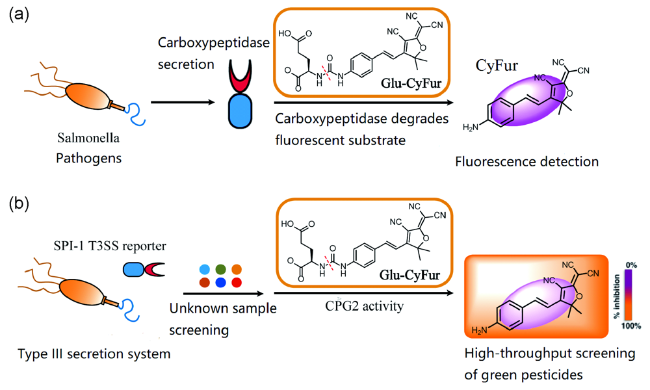
4 Type Ⅲ secretion system-targeted small-molecule fluorescent probes
5 γ-Aminobutyric acid receptor-targeted small-molecule fluorescent pesticides
6 Acetohydroxyacid synthase-targeted small-molecule fluorescent probes
7 Conclusion and outlook
| [1] |
Huang J S, Guo K X, You L J. South Chin. Agric., 2019, 13(14):165.
(黄佳盛, 郭凯先, 尤李俊. 南方农业, 2019, 13(14): 165.)
|
| [2] |
Liu Y R, Gao R F, Yang J W, Han L Y, Qi X F, Mao M M. J. Food Saf. Qual., 2019, 10(24):8562.
(刘艳容, 高瑞峰, 杨佳玮, 韩璐瑶, 祁雪峰, 毛敏明. 食品安全质量检测学报, 2019, 10(24): 8562.)
|
| [3] |
An H B, Li Z S. Appl. Sci. Technol., 2003, 30(9):47.
(安红波, 李占双. 应用科技, 2003, 30(9): 47.)
|
| [4] |
Shao X S, Du S Q, Li Z, Qian X H. World Pestic., 2020, 42(4):16.
(邵旭升, 杜少卿, 李忠, 钱旭红. 世界农药, 2020, 42(4): 16.)
|
| [5] |
Croston G E. Expert. Opin. Drug Discov., 2017, 12(5):427.
|
| [6] |
Meissner G. J. Gen. Physiol., 2017, 149(12):1065.
|
| [7] |
Kato K, Kiyonaka S, Sawaguchi Y, Tohnishi M, Masaki T, Yasokawa N, Mizuno Y, Mori E, Inoue K, Hamachi I, Takeshima H, Mori Y S. Biochemistry, 2009, 48(43):10342.
DOI: 10.1021/bi900866s |
| [8] |
Sun L N, Qiu G S, Cui L, Ma C S, Yuan H Z. Pestic. Biochem. Physiol., 2015, 123:56.
|
| [9] |
Guo L, Liang P, Fang K, Chu D. Pestic. Biochem. Physiol., 2017, 142:162.
|
| [10] |
Tan H J. World Pestic., 2019, 41(5):60.
(谭海军. 世界农药, 2019, 41(5):60.)
|
| [11] |
Xing J H, Zhu B C, Yuan J, Yu J P, Dong D Z, Zhou D Y, Hu D S, Chen J. Chin. J. Pestic. Sci., 2013, 15(2):159.
(邢家华, 朱冰春, 袁静, 郁季平, 董德臻, 周冬英, 胡冬松, 陈杰. 农药学学报, 2013, 15(2):159.)
|
| [12] |
Tan H J. Fine Spec. Chem., 2020, 28(2):31.
(谭海军. 精细与专用化学品. 2020, 28(2):31.)
|
| [13] |
Mueller K H. US9512144, 2016.
|
| [14] |
Nakao T, Banba S. Bioorg. Med. Chem., 2016, 24(3):372.
|
| [15] |
Katsuta H, Nomura M, Wakita T, Daido H, Kobayashi Y, Kawahara A, Banba S. J. Pestic. Sci., 2019, 44(2):120.
|
| [16] |
Bo Y L. Pestic. Market News, 2018, 20:27.
(柏亚罗. 农药市场信息, 2018, 20:27.).
|
| [17] |
Sun Y, Xu L, Chen Q, Qin W J, Huang S J, Jiang Y, Qin H G. Pest. Manag. Sci, 2018, 74(6):1416.
|
| [18] |
Selby T P, Lahm G P, Stevenson T M. Pest. Manag. Sci., 2017, 73(4):658.
|
| [19] |
Li B, Yang H B, Wang J F, Song Y Q. Mod. Agrochem., 2014,(3):17.
(李斌, 杨辉斌, 王军锋, 宋玉泉. 现代农药, 2014,(3):17.)
|
| [20] |
He X L. World Pestic., 2016, 38(3):60.
(何秀玲. 世界农药, 2016, 38(3):60.)
|
| [21] |
Sakuma Haruhiko. CN 101528040, 2009.
|
| [22] |
Wang Y, Guo L, Qi S Z, Zhang H, Liu K C, Liu R Q, Liang P, Casida J, Liu S Z. Molecules, 2014, 19(4):4105.
DOI: 10.3390/molecules19044105 PMID: 24699151
Diamide insecticides with high efficacy against pests and good environmental safety are broadly applied in crop protection. They act at a poorly-defined site in the very complex ryanodine (Ry) receptor (RyR) potentially accessible to a fluorescent probe. Two N-propynyl analogs of the major anthranilic diamide insecticides chlorantraniliprole (Chlo) and cyantraniliprole (Cyan) were accordingly synthesized and converted into two fluorescent ligands by click reaction coupling with 3-azido-7-hydroxy-2H-chromen-2-one. The new diamide analogs and fluorescent ligands were shown to be nearly as potent as Chlo and Cyan in inhibition of [3H]Chlo binding and stimulation of [3H]Ry binding in house fly thoracic muscle RyR. Although the newly synthesized compounds had only moderate activity in insect larvicidal activity assays, their high in vitro potency in a validated insect RyR binding assay encourages further development of fluorescent probes for insect RyRs.
|
| [23] |
Chen J, Xue L, Wei R S, Liu S Z, Yin C C. Biochem. Biophys. Res. Commun., 2019, 508(2):633.
|
| [24] |
Liu T F, Yang D F, Deng J H, Dong M H. Chin. Agric. Sci. Bull., 2015, 31(3):221.
(刘腾飞, 杨代凤, 邓金花, 董明辉. 中国农学通报, 2015, 31(3):221.)
|
| [25] |
Qi H L, Cui L, Wang Q Q, Liu F, Rui C H. Plant Prot., 2017, 43(1):112.
(齐浩亮, 崔丽, 王芹芹, 刘峰, 芮昌辉. 植物保护, 2017, 43(1):112.)
|
| [26] |
Isaacs A K, Qi S Z, Sarpong R, Casida J E. Chem. Res. Toxicol., 2012, 25(8):1571.
DOI: 10.1021/tx300326m PMID: 22856329
Radiolabeled anthranilic diamide insecticide [N-C(3)H(3)]chlorantraniliprole was synthesized at high specific activity. It was compared with phthalic diamide insecticide flubendiamide and [(3)H]ryanodine in radioligand binding studies with house fly muscle membranes to provide the first direct evidence with a native insect ryanodine receptor that the major anthranilic and phthalic diamide insecticides bind at different allosterically coupled sites, i.e., there are three distinct Ca(2+)-release channel targets for insecticide action.
|
| [27] |
Qi S Z, Lümmen P, Nauen R, Casida J E. J. Agric. Food Chem., 2014, 62(18):4077.
DOI: 10.1021/jf501236h |
| [28] |
Li Z Z, Jiang H, Liu S N, Li Y X, Yuchi Z G, Gao Q Z. Anal. Chimica Acta, 2020, 1108:108.
|
| [29] |
Liao M, Li Q B, Yang Z K, Feng T, Xu Z Y, Liu Q, Liu S Z. Ann. N.Y. Acad. Sci., 2020, 1475(1):43.
DOI: 10.1111/nyas.14362 PMID: 32483859
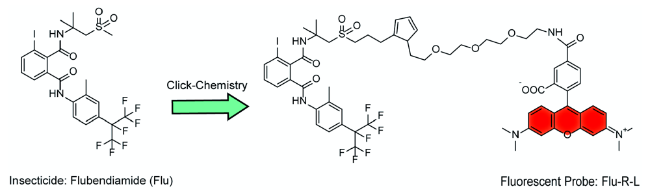
There is a significant need to study the binding of active compounds to the specific sites on insect ryanodine receptors (RyRs) that are the targets of two novel classes of diamide insecticides to which insects are becoming increasingly resistant. Here, we describe a rapid assay to study the action of potential compounds on the flubendiamide (Flu) binding site of insect RyRs that uses a fluorescence polarization assay with the fluorescence probe Flu-R-L that we synthesized. The IC of Flu for inhibiting probe binding on insect RyR was 18.82 ng/mL. The binding of 86 novel phthalic diamide derivatives on insect RyRs was studied using this newly established assay, and the compounds that exhibited high-affinity binding in the assay also possessed in vivo insecticidal activity against Plutella xylostella. Thus, Flu-R-L is a highly selective and sensitive fluorescence probe for studying the binding affinity of novel compounds to the Flu binding site of insect RyRs. The assay based on Flu-R-L is a rapid, accurate, and sensitive method for the screening of potentially bioactive molecules that bind specifically to insect RyRs. © 2020 New York Academy of Sciences.
|
| [30] |
Chowdhury R, Candela-Lena J I, Chan M C, Greenald D J, Yeoh K K, Tian Y M, McDonough M A, Tumber A, Rose N R, Conejo-Garcia A, Demetriades M, Mathavan S, Kawamura A, Lee M K, van Eeden F, Pugh C W, Ratcliffe P J, Schofield C J. ACS Chem. Biol., 2013, 8(7):1488.
DOI: 10.1021/cb400088q |
| [31] |
Vachal P, Miao S W, Pierce J M, Guiadeen D, Colandrea V J, Wyvratt M J, Salowe S P, Sonatore L M, Milligan J A, Hajdu R, Gollapudi A, Keohane C A, Lingham R B, Mandala S M, DeMartino J A, Tong X C, Wolff M, Steinhuebel D, Kieczykowski G R, Fleitz F J, Chapman K, Athanasopoulos J, Adam G, Akyuz C D, Jena D K, Lusen J W, Meng J C, Stein B D, Xia L, Sherer E C, Hale J J. J. Med. Chem., 2012, 55(7):2945.
DOI: 10.1021/jm201542d PMID: 22364528
The discovery of 1,3,8-triazaspiro[4.5]decane-2,4-diones (spirohydantoins) as a structural class of pan-inhibitors of the prolyl hydroxylase (PHD) family of enzymes for the treatment of anemia is described. The initial hit class, spirooxindoles, was identified through affinity selection mass spectrometry (AS-MS) and optimized for PHD2 inhibition and optimal PK/PD profile (short-acting PHDi inhibitors). 1,3,8-Triazaspiro[4.5]decane-2,4-diones (spirohydantoins) were optimized as an advanced lead class derived from the original spiroindole hit. A new set of general conditions for C-N coupling, developed using a high-throughput experimentation (HTE) technique, enabled a full SAR analysis of the spirohydantoins. This rapid and directed SAR exploration has resulted in the first reported examples of hydantoin derivatives with good PK in preclinical species. Potassium channel off-target activity (hERG) was successfully eliminated through the systematic introduction of acidic functionality to the molecular structure. Undesired upregulation of alanine aminotransferese (ALT) liver enzymes was mitigated and a robust on-/off-target margin was achieved. Spirohydantoins represent a class of highly efficacious, short-acting PHD1-3 inhibitors causing a robust erythropoietin (EPO) upregulation in vivo in multiple preclinical species. This profile deems spirohydantoins as attractive short-acting PHDi inhibitors with the potential for treatment of anemia. © 2012 American Chemical Society
|
| [32] |
Li T C, Chen J, Fan X B, Chen W W, Zhang W Q. Pest. Manag. Sci., 2017, 73(7):1529.
|
| [33] |
Zimoch L, Hogenkamp D G, Kramer K J, Muthukrishnan S, Merzendorfer H. Insect Biochem. Mol. Biol., 2005, 35(6):515.
|
| [34] |
Batran R Z, Khedr M A, Abdel Latif N A, Abd El Aty A A, Shehata A N. J. Mol. Struct., 2019, 1180:260.
DOI: 10.1016/j.molstruc.2018.11.099
The inhibition of chitinase activity is considered of great importance for the development of novel antifungal agents. Here we explore 4-hydroxycoumarins as a novel natural product-derived scaffold for inhibiting chitinases. A new series of 4-hydoxycoumarin derivatives containing Schiffs base motif in the 3 position was synthesized and computationally predicted for chitinase binding affinity. Docking simulation study was carried out using the built homology models of specified fungal species and the top ranked molecules with promising binding affinity were subjected to further molecular dynamic simulation to evaluate their binding stability. The top compounds were then tested in vitro against three phytopathogenic fungi, including Fusarium solani, Fusarium oxysporium and Aspergillus niger in addition to three candida species including C albicans, C. tropicalis and C krusei. The active antifungal candidates were further assessed for chitinase inhibition effect. Most of the tested compounds displayed promising antifungal effects. The 2,4-dichlorophenyl Schiffs base 2e exhibited a wide range of antifungal activity against most of the tested fungi and yeast while the 3-fluorophenyl Schiff's base 2a showed the highest anti-candidal effects. Both Schiffs bases 2a and 2e displayed the highest chitinase inhibition effects (IC50=1.0 mM) having the same potential as the previously described chitinase inhibitor CI-4 (IC50=1.2 mM). The target compounds have high ligand efficiency and binding stability, and as such are promising leads for future development of chitinase inhibitors. (C) 2018 Elsevier B.V.
|
| [35] |
Li B, Wang K Y, Zhang R, Li B H, Shen Y L, Ji Q G. Eur. J. Med. Chem., 2019, 182:111669.
|
| [36] |
Zhang P, Zhao Y H, Wang Q H, Mu W, Liu F. Pestic. Biochem. Physiol., 2017, 136:80.
DOI: S0048-3575(16)30076-1 PMID: 28187835
Bradysia odoriphaga Yang and Zhang is the primary insect pest that affects Chinese chive in northern China. Nevertheless, very few studies have been conducted on the use of chitin synthesis inhibitors (CSIs) for the control of B. odoriphaga. Here, lethal and sublethal effects of the CSI chlorfluazuron on B. odoriphaga were studied to explore the use for integrated pest management (IPM) of B. odoriphaga. A contact and ingestion toxicity bioassay showed that chlorfluazuron was more active against B. odoriphaga than three other CSIs, with a 72h LC of 0.1593mg/L. Treatment with the LC dose of chlorfluazuron decreased both the intrinsic and finite rates of increase of B. odoriphaga, in addition to reproduction rate, survival rate, and fecundity, and the mean generation time, total preovipositional period and larval development duration were shortened, compared with those of the control and LC groups. The mean generation time, total preovipositional period and larval development duration were all also markedly decreased by treatment with chlorfluazuron at the LC. Furthermore, chlorfluazuron inhibited the feeding of the final instar larvae for a short period. Glutathione S-transferase and microsomal mixed function oxidase activities increased after exposure to the chemical. These results showed that chlorfluazuron at the sublethal LC treatment inhibited B. odoriphaga population growth, whereas the danger of causing rapid population growth by using a lower sublethal concentration was demonstrated with the sublethal LC treatment. Therefore, chlorfluazuron should be used with caution in an IPM program for B. odoriphaga. Copyright © 2016 Elsevier Inc. All rights reserved.
|
| [37] |
Cohen E. Annu. Rev. Entomol., 1987, 32(1):71.
|
| [38] |
Bahmed K, Quilès F, Wathier M, Bonaly R, Benallaoua S, Pucci B, Coulon J. Process. Biochem., 2005, 40(7):2523.
|
| [39] |
Chen Q, Zhang J W, Chen L L, Yang J, Yang X L, Ling Y, Yang Q. Chin. Chem. Lett., 2017, 28(6):1232.
|
| [40] |
Goedken E R, O’Brien R F, Xiang T, Banach D L, Marchie S C, Barlow E H, Hubbard S, Mankovich J A, Jiang J J, Richardson P L, Cuff C A, Cherniack A D. Protein Expr. Purif., 2011, 75(1):55.
|
| [41] |
Dong L L, Shen S Q, Lu H Z, Jin S H, Zhang J J. ACS Sens., 2019, 4(5):1222.
|
| [42] |
Li T H, Zhang D D, Oo T N, San M M, Mon A M, Hein P P, Wang Y H, Lu C H, Yang X F. Evid. Based Complementary Altern. Med., 2018,2018: 1.
|
| [43] |
Roca I, Akova M, Baquero F, Carlet J, Cavaleri M, Coenen S, Cohen J, Findlay D, Gyssens I, Heure O E, Kahlmeter G, Kruse H, Laxminarayan R, Liébana E, López-Cerero L, MacGowan A, Martins M, Rodríguez-Baño J, Rolain J M, Segovia C, Sigauque B, Tacconelli E, Wellington E, Vila J. New Microbes New Infect., 2015, 6:22.
|
| [44] |
Schesser Bartra S, Lorica C, Qian L F, Gong X, Bahnan W, Barreras H Jr, Hernandez R, Li Z W, Plano G V, Schesser K. Front. Cell. Infect. Microbiol., 2019, 9:23.
|
| [45] |
Fan S S, Tian F, Li J Y, Hutchins W, Chen H M, Yang F H, Yuan X C, Cui Z N, Yang C H, He C Y. Mol. Plant Pathol., 2017, 18(4):555.
|
| [46] |
Xiang X W, Tao H, Jiang S, Zhang L H, Cui Z N. Pestic. Biochem. Physiol., 2018, 149:89.
|
| [47] |
Tao H, Tian H, Jiang S, Xiang X W, Lin Y N, Ahmed W, Tang R Y, Cui Z N. Pestic. Biochem. Physiol., 2019, 160:87.
|
| [48] |
Pendergrass H A, May A E. Antibiotics, 2019, 8(4):162.
|
| [49] |
Lan N H, Su D S, Zhou L, Qin X, Huang P F, Yang M, Jiang B L. Acta. Phytopathologica. Sinica., 2019, 49(2):262.
(蓝逆寒, 苏德山, 周乐, 覃霞, 黄佩芳, 杨梅, 姜伯乐. 植物病理学报, 2019, 49(2):262.)
|
| [50] |
Allombert J, Vianney A, Charpentier X. Methods in Molecular Biology. New York: Springer New York, 2017.489.
|
| [51] |
Yount J S, Tsou L K, Dossa P D, Kullas A L, van der Velden A W M, Hang H C. J. Am. Chem. Soc., 2010, 132(24):8244.
DOI: 10.1021/ja102257v |
| [52] |
Tsou L K, Lara-Tejero M, RoseFigura J, Zhang Z J, Wang Y C, Yount J S, Lefebre M, Dossa P D, Kato J, Guan F L, Lam W, Cheng Y C, Galán J E, Hang H C. J. Am. Chem. Soc., 2016, 138(7):2209.
DOI: 10.1021/jacs.5b11575 PMID: 26847396
Traditional Chinese Medicines (TCMs) have been historically used to treat bacterial infections. However, the molecules responsible for these anti-infective properties and their potential mechanisms of action have remained elusive. Using a high-throughput assay for type III protein secretion in Salmonella enterica serovar Typhimurium, we discovered that several TCMs can attenuate this key virulence pathway without affecting bacterial growth. Among the active TCMs, we discovered that baicalein, a specific flavonoid from Scutellaria baicalensis, targets S. Typhimurium pathogenicity island-1 (SPI-1) type III secretion system (T3SS) effectors and translocases to inhibit bacterial invasion of epithelial cells. Structurally related flavonoids present in other TCMs, such as quercetin, also inactivated the SPI-1 T3SS and attenuated S. Typhimurium invasion. Our results demonstrate that specific plant metabolites from TCMs can directly interfere with key bacterial virulence pathways and reveal a previously unappreciated mechanism of action for anti-infective medicinal plants.
|
| [53] |
Tsou L K, Yount J S, Hang H C. Bioorg. Med. Chem., 2017, 25(11):2883.
|
| [54] |
Büttner D, Bonas U. Curr. Opin. Microbiol., 2006, 9(2):193.
|
| [55] |
Wei Q, Wu S F, Gao C F. Agrochemicals, 2014, 53(12):859.
(魏琪, 吴顺凡, 高聪芬. 农药, 2014, 53(12):859.)
|
| [56] |
Gibbons D, Morrissey C, Mineau P. Environ. Sci. Pollut. Res., 2015, 22(1):103.
|
| [57] |
Mahler B J, van Metre P C, Wilson J T, Musgrove M, Zaugg S D, Burkhardt M R. Environ. Sci. Technol., 2009, 43(15):5665.
DOI: 10.1021/es901292a |
| [58] |
Dedryver C A, Ralec A L, Fabreb F. C. R. Biol., 2010, 333(6):539.
|
| [59] |
Wang F, Meng X H, Wang H. Agric. Sci., 2014, 4(5):99.
(王峰, 孟祥鹤, 王菡. 农业科学, 2014, 4(5):99.)
|
| [60] |
Wu Z D, Du Y T, Zhou Q, Chen L Q. Pestic. Biochem. Physiol., 2020, 163:51.
|
| [61] |
Garcia M D, Wang J G, Lonhienne T, Guddat L W. FEBS J., 2017, 284(13):2037.
|
| [62] |
Chen W, Wei W, Zhou S, Li Y H, Zhang X, Tong J, Li Y X, Li Z M. Chem. J. Chinese Univ., 2015, 36(4):672.
DOI: 10.1002/cjoc.v36.7 |
| [63] |
Wang J G. Chin. J. Pestic. Sci., 2014, 16(4):367.
|
| [64] |
Eoyang L, Silverman P M. J. Bacteriol., 1984, 157(1):184.
PMID: 6360995
Acetohydroxyacid synthase I from Escherichia coli K-12 has been purified to near homogeneity. Analysis of the purified enzyme by sodium dodecyl sulfate-polyacrylamide gel electrophoresis showed the presence of two polypeptides, one with a molecular weight of 60,000 and one with a molecular weight of 9,500. These two polypeptides were present in constant proportion to each other and to enzyme activity. The molar ratio of the two polypeptides (Mr 9,500:60,000), estimated from stained polyacrylamide gels, was 1. Antisera raised against the 60,000 Mr polypeptide precipitated both the 60,000 and the 9,500 Mr polypeptides from extracts of cells labeled with [35S]methionine. The addition of sodium dodecyl sulfate before immunoprecipitation eliminated the smaller polypeptide, and only the larger one was recovered. The hydrodynamic properties of the native enzyme confirmed a previous report that the largest enzymatically active species has a molecular weight of about 200,000; this species contains both the 60,000- and 9,500-molecular-weight polypeptides.
|
| [65] |
Hattori J, Rutledge R, Labbé H, Brown D, Sunohara G, Miki B. Mol. Gen. Genet. MGG, 1992, 232(2):167.
DOI: 10.1007/BF00279993 |
| [66] |
Chang S I, Kang M K, Choi J D, Namgoong S K. Biochem. Biophys. Res. Commun., 1997, 234(3):549.
|
| [67] |
Arabet D, Tempel S, Fons M, Denis Y, Jourlin-Castelli C, Armitano J, Redelberger D, Iobbi-Nivol C, Boulahrouf A, Méjean V. Environ. Sci. Pollut. Res., 2014, 21(8):5619.
|
| [68] |
Mao D L, Michelmore S, Paull J, Preston C, Sutton T, Oldach K, Yang S Y, McMurray L. Pest. Manag. Sci., 2019, 75(10):2698.
DOI: 10.1002/ps.v75.10 |
| [69] |
Liu Y C, Qu R Y, Chen Q, Yang J F, Niu C W, Zhen X, Yang G F. J. Agric. Food Chem., 2016, 64(24):4845.
|
| [70] |
Wang W, Wang Y C, Li Z, Wang H Y, Yu Z Y, Lu L, Ye Q F. Sci. Total. Environ., 2014, 472:582.
|
| [71] |
Chen G F, Qiao Y X, Zhang X B, Liu F, Liao H, Zhang R Y, Dong J N, Tao B. Bull. Environ. Contam. Toxicol., 2019, 102(6):854.
|
| [72] |
Zhong M M, Wang T L, Hu J Y. Environ. Monit. Assess., 2015, 187(6):1.
|
| [73] |
Garcia M D, Nouwens A, Lonhienne T G, Guddata L W. Proc. Natl. Acad. Sci. U.S.A., 2017, 114(7):E1091.
|
| [74] |
Duggleby R G, Pang S S, Yu H Q, Guddat L W. Eur. J. Biochem., 2003, 270(13):2895.
PMID: 12823560
Acetohydroxyacid synthase (AHAS, EC 4.1.3.18) catalyses the first step in branched-chain amino acid biosynthesis and is the target for sulfonylurea and imidazolinone herbicides, which act as potent and specific inhibitors. Mutants of the enzyme have been identified that are resistant to particular herbicides. However, the selectivity of these mutants towards various sulfonylureas and imidazolinones has not been determined systematically. Now that the structure of the yeast enzyme is known, both in the absence and presence of a bound herbicide, a detailed understanding of the molecular interactions between the enzyme and its inhibitors becomes possible. Here we construct 10 active mutants of yeast AHAS, purify the enzymes and determine their sensitivity to six sulfonylureas and three imidazolinones. An additional three active mutants were constructed with a view to increasing imidazolinone sensitivity. These three variants were purified and tested for their sensitivity to the imidazolinones only. Substantial differences are observed in the sensitivity of the 13 mutants to the various inhibitors and these differences are interpreted in terms of the structure of the herbicide-binding site on the enzyme.
|
| [75] |
Riar D S, Tehranchian P, Norsworthy J K, Nandula V, McElroy S, Srivastava V, Chen S, Bond J A, Scott R C. Weed Sci., 2015, 63(4):748.
|
| [76] |
Yu Q, Powles S B. Pest. Manag. Sci., 2014, 70(9):1340.
DOI: 10.1002/ps.3710 |
| [77] |
Kreiner J M, Stinchcombe J R, Wright S I. Annu. Rev. Plant Biol., 2018, 69(1):611.
|
| [78] |
Singh B K, Stidham M A, Shaner D L. Anal. Biochem., 1988, 171(1):173.
PMID: 3407914
Acetohydroxyacid synthase (AHAS), also known as acetolactate synthase, has received attention recently because of the finding that it is the site of action of several new herbicides. The most commonly used assay for detecting the enzyme is spectrophotometric involving an indirect detection of the product acetolactate. The assay involves the conversion of the end product acetolactate to acetoin and the detection of acetoin via the formation of a creatine and naphthol complex. There is considerable variability in the literature as to the details of this assay. We have investigated a number of factors involved in detecting AHAS in crude ammonium sulfate precipitates using this spectrophotometric method. Substrate and cofactor saturation levels, pH optimum, and temperature optimum have been determined. We have also optimized a number of factors involved in the generation and the detection of acetoin from acetolactate. The results of these experiments can serve as a reference for new investigators in the study of AHAS.
|
| [79] |
Schloss J V, van Dyk D E, Vasta J F, Kutny R M. Biochemistry, 1985, 24(18):4952.
PMID: 3907697
A facile purification has been devised for recombinantly produced Salmonella typhimurium acetolactate synthase isozyme II. Purification of the enzyme was made possible by determining the complex set of factors that lead to loss of enzymic activity with this rather labile enzyme. When complexed with thiamin pyrophosphate, FAD, and magnesium, acetolactate synthase is subject to oxygen-dependent inactivation, a property not shared by the enzyme-FAD complex. When divorced from all of its tightly bound cofactors, losses of the enzymic activity are encountered at low ionic strength, especially at low protein concentrations. If purified and stored as the enzyme-FAD complex, acetolactate synthase is quite stable. The enzyme is composed of two types of subunits, a result that was not anticipated from previous studies of ilvG (the gene that codes for the large subunit of acetolactate synthase). These subunits were determined to be in equal molar ratio in the purified enzyme from the distribution of radioactivity between the two subunits after carboxymethylation with iodo[14C]acetate and their respective amino acid compositions. Besides the expected ilvG gene product (59.3 kDa), purified acetolactate synthase contained a smaller subunit (9.7 kDa; designated here as the ilvM gene product). On the basis of sequence homology of the small subunit with that coded for by the corresponding Escherichia coli gene sequence [Lawther, R. P., Calhoun, D. H., Adams, C. W., Hauser, C. A., Gray, J., & Hatfield, G. W. (1981) Proc. Natl. Acad. Sci. U.S.A. 78, 922-925], it is encoded by the region between ilvG and ilvE, beginning at base-pair (bp) 1914 (relative to the point of transcription initiation).(ABSTRACT TRUNCATED AT 250 WORDS)
|
| [80] |
Xie Y H, Zhang C Y, Wang Z H, Wei C, Liao N J, Wen X, Niu C W, Yi L, Wang Z J, Xi Z. Anal. Chem., 2019, 91(21):13582.
|
/
| 〈 |
|
〉 |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}