




1 引言
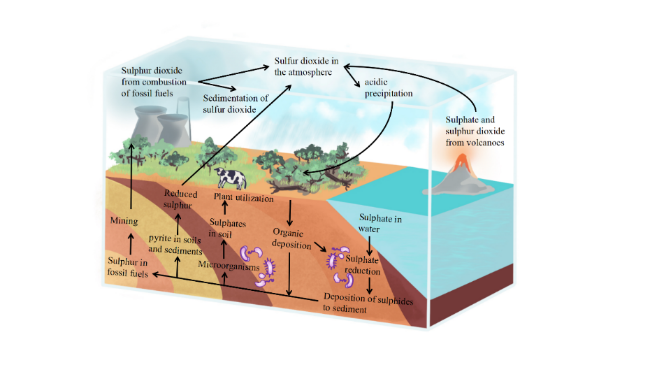
硫酸盐还原菌(sulfate-reducing bacteria,SRB)作为一类广泛存在的厌氧菌群,如图1所示SRB广泛分布于土壤、海洋、油井、排水和动物肠道等生态系统中,其在全球硫碳氮循环中起到重要作用。
SRB具备利用多种电子供体和电子受体的能力,使其在废水处理过程中表现出了独特的优势,从而在不同类型的废水处理中都有广泛应用。例如,采用UASB反应器在35 ℃厌氧条件下可有效处理常规冲厕黑水,研究发现反应器中的优势菌群为硫酸盐还原菌,而在该体系下氢营养型产甲烷菌活性较低。利用SRB对印染废水中偶氮染料(procion red HE-7B)进行脱除和降解,在碱性条件下(pH=11)和高COD下,HE-7B的去除率达90%以上。在重金属废水处理中,SRB通过硫酸盐还原促使了金属硫化物的生成,从而对废水中的重金属进行有效的去除。
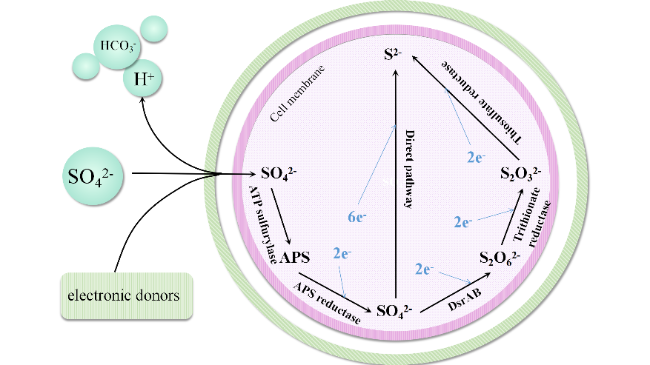
SRB还原硫酸盐可能主要有两种途径[1]。一种是同化硫酸盐还原,SRB利用硫酸盐还原产物直接合成细胞材料。另一种途径是异化硫酸盐还原,SRB可以将有机碳源完全氧化分解为二氧化碳,或者在厌氧条件下不完全氧化分解为乙酸。同时,底物水平磷酸化产生少量三磷酸腺苷(adenosine triphosphate, ATP)和高能电子;磷酸化释放的高能电子通过电子传递链(黄素蛋白、细胞色素c等)分步传递给SRB,SRB则通过“电子传递磷酸化”产生大量ATP,为生化过程提供能量。最后,硫氧化物通过消耗ATP提供能量被还原为S2−。
因此,利用自养型SRB来进行硫酸盐还原既减少了有机质的投加,又降低了MB的竞争反应。但自养型SRB处理硫酸盐并不能适用于所有类型的污废水:在绝大部分的含硫酸盐废水中,平衡SRB和MB的常见方法是控制环境因素,为SRB提供优势的生长条件,但对于水质波动较大的废水和含有复杂成分的污水,这种平衡方法也存在着局限性。
为了在碳源利用和菌群竞争之间寻求平衡,引入了电偶合技术。近年来,电耦合硫酸盐去除技术在含硫酸盐废水处理中得到了应用。面对环境中有限的电子供体,硫酸盐还原菌不可避免地会与环境中的其他菌群进行竞争或共生。根据硫酸盐的浓度变化,硫酸盐还原菌所进行的主要电子转移途径也会发生变化,电子供体的竞争模式也会随之发生变化,而反应体系中电场的加入将会对自然竞争结果产生显著影响。外加电场对体系中菌群的电子传递过程进行干预,利用调节菌群竞争过程从而实现提高硫酸盐去除效能的目的。例如,微生物电化学系统技术(microbial electrochemical system technology,MES)可以通过生物阴极的构建原位产生H2,即通过电介导过程对SRB和MB的竞争过程产生影响。此外,在MES的典型工艺微生物燃料电池(microbial fuel cell, MFC)中,有机化合物或单质硫在阳极室中被氧化,电子则通过外导线释放转移到阴极,并被微生物作为电子供体消耗[3]。由于微生物具有着直接或间接利用来自阴极的电子作为电子供体还原污染物(如硫酸盐、硝酸盐、Cr6+和苯胺等)[4⇓~6]的能力,在含有多类污染物的废水中设置微生物电化学系统,可以同时去除在热力学上不相近的两种或多种污染物的成为可能。
SRB在污废水处理中扮演着至关重要的角色,其在含硫酸盐废水以及含硫化物废水处理中都具有独特优势。此外,SRB与其他微生物之间的相互作用将影响整个微生物群落的结构和功能。通过研究SRB在菌群竞争中的地位和作用,可以更好地理解微生物群落的演替规律和稳定机制,更有利于指导废水处理工艺的设计和优化。本文主要综述了关于硫酸盐还原菌在菌群竞争和电子供体利用机理方面的研究与成果,特别对微生物电化学系统影响下的SRB菌群活动进行了归纳,并对其进行了整理、分析阐述和扩展,客观地提出了SRB与其他菌群竞争的影响因素,以便进一步探索研究控制方法,为提高含硫废水生化处理水平提供相关帮助。
2 硫酸盐还原菌在水环境中的菌群关系
在纸浆、印染、酿造等行业废水和酸性矿山废水中,硫酸盐浓度可达到数百至上千毫克升,这对传统有机物污染去除过程产生严重影响。在富含硫酸盐的环境中, SRB在有机物氧化过程中使用硫酸盐作为终端电子受体,从而产生硫化物,其间,SRB将会与体系中的其他菌群发生竞争。而在缺乏硫酸盐的环境中SRB和MB在有机物降解过程中不是竞争关系而是共生关系。由此可见,通过对SRB在水处理过程中与不同菌群间的相互作用研究,可以获知SRB与其他菌群共生与竞争的影响因素,从微生物组学角度揭示SRB在微生物种群中的调控机理,为优化其应用于水处理工艺提供微生物学理论基础。
2.1 共生关系
Reeburgh[7]在1976年提出,甲烷的厌氧氧化可以和硫酸盐还原相结合,随后,Bryan等[8]在厌氧产甲烷的反应器中发现硫酸盐还原菌可以作为产丙酮菌存在。之后的大量研究也揭示了硫酸盐依赖甲烷氧化的微生物学过程。与此同时,在缺乏硫酸盐的自然水体中,SRB在有机物发酵和厌氧氧化中也起到了重要作用。因此,可以认为在绝大多数缺乏硫酸盐的环境中SRB和MB对有机物的降解是在共生关系下进行的而不是竞争关系。共生细菌(syntrophobacter species)是一类特殊的硫酸盐还原菌,可以在丙酸盐和硫酸盐环境中生长,在与氢营养型产甲烷菌共培养时,能够将丙酸盐转化为醋酸盐、二氧化碳和氢。如脱硫弧菌(Desulfovibrio)和脱硫杆菌(Desulfomicroium)可通过利用发酵丙酮酸生成的乙酸、二氧化碳和氢气来进行代谢生长。在氢营养型产甲烷菌将氢利用完后,SRB能将乳酸盐和乙醇氧化为醋酸盐作为生长基质[8]。脱硫棒菌(Desulfobulbus)同样可以利用丙酸盐和硫酸盐进行生长,但与共生菌群不同的是,在与产甲烷菌共培养时其无法将丙酸盐转化为乙酸盐,而在无硫酸盐的环境中,Desulfobulbus可以将乳酸和乙醇(二氧化碳存在时)发酵成醋酸盐和丙酸盐。
在硫酸盐环境中,SRB的富集可以通过改变代谢途径和电子传递来促进甲烷产生。Zan等[9]在食物垃圾和废物活性污泥共消化研究中,通过往体系中添加硫酸盐,使SRB在共消化中作为产乙酸菌将丙酸转化为乙酸,从热力学角度来看,这为MB提供了替代代谢途径,从而可以通过调控硫酸盐浓度提高厌氧共消化中的产甲烷效率。Li等 [10] 采用两相厌氧反应器(COD/SO42-:3)中的产酸相(产酸-硫酸盐还原反应)进行硫酸盐还原,从而避免SRB与MB的竞争,有效提高含硫酸盐有机废水的去除效率。其中SRB细菌利用水解酸化细菌的液相末端代谢物作为电子供体,利用SO42-作为电子受体进行硫酸盐还原,SRB菌的存在可以促进产酸菌的代谢率。在自然竞争过程中,硫酸盐的存在及浓度的高低对竞争的结果具有重要的影响,SRB与MB的共生关系更多地存在于硫酸盐浓度不高的环境中。
在自然体系下,SRB可以与多种氧化菌和脱氢菌共生,形成菌群后共同完成生态系统中的循环过程。与SRB共生的氧化菌包括铁氧化菌、锰氧化菌等,它们可以利用SRB产生的硫化氢气体作为电子供体,完成自身的代谢过程。同时,SRB也可以与脱氢菌共生,如甲烷菌等,可以利用SRB产生的甲烷气体作为电子供体。然而,当环境中存在硫酸盐而有机底物浓度较低时,硫酸盐还原菌就会与其他菌群竞争资源,这种竞争将会抑制其他菌群的生长,从而使SRB在生态系统中占据优势地位。
2.2 竞争关系
硫酸盐还原菌作为一类厌氧环境中常见的以异养为主的微生物,不论是硫酸盐还原菌之间,还是硫酸盐还原菌与其他厌氧菌群之间都会存在着竞争行为,竞争的主要资源包括硫酸盐和有机物。在竞争中,SRB会通过多种方式来争夺资源。首先,SRB可以通过分泌特定的酶来降解有机物,从而获得更多的营养物质。其次,SRB可能会产生抗生素等化合物,以抑制其他菌群的生长,从而减少竞争。此外,一些SRB还可以形成有利于自身附着的生物膜,以占据更多的生态优势。不仅如此,硫酸盐还原菌作为一类菌属,不同的硫酸盐还原菌也会体现出不同的营养模式,这便对彼此的生存和代谢造成影响。根据碳代谢途径SRB可分为两大类:一类能将有机物完全降解为CO2;另一类则进行不完全氧化将有机物转化为乙酸盐。因此SRB不仅与MB竞争H2/CO2和醋酸盐,也与合成细菌竞争底物,如丙酸盐、丁酸盐和乙醇等,而这些是厌氧消化过程中的关键中间体。对底物的竞争同时体现在对有机物的降解途径上,硫酸盐还原的环境(图3a)和产甲烷环境(图3b)中的有机物降解是不同的[11]。由于MB能直接利用的底物主要是CO2、H2和乙酸盐、,而在以有机酸,如乳酸、丙酸和丁酸作为生长底物时,SRB更具有优势,且SRB在可利用底物种类上也更为丰富。因此在硫酸盐存在的环境中,SRB能直接利用更多的底物进行代谢生长。而在产甲烷反应器中,化合物需要先被丙酮菌和产甲烷菌的共生群落分解后才能形成产甲烷菌的营养底物(图3b)。
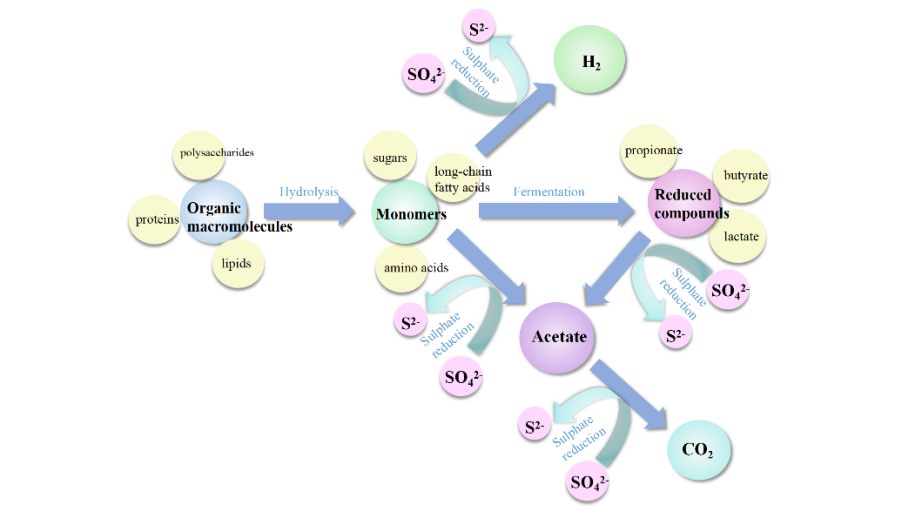
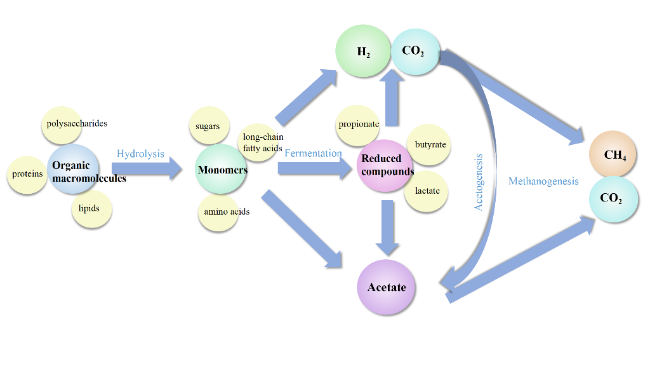
在富含硫酸盐废水的厌氧处理过程中,通常会存在硫化物毒性危害、甲烷产量低、H2S等有毒气体生成和腐蚀等问题。其中,H2S的产生会对体系中的菌群造成严重的毒害作用从而抑制反应器性能的提高,通过对SRB和MB之间的竞争调节可以缓解并平衡这些现象。研究证实,随着SRB生物量的增加,产甲烷古菌(methanogenic archaea,MA)的甲烷产率和生物量逐渐下降,这表明SRB可以在低乙酸浓度下与MA进行有效的竞争[12]。从热力学的角度来看,SRB比MA和产丙酮细菌(AcB)更有竞争力。然而,Jing等[13]研究发现在运行180天处理碳硫比(COD/SO42-)比为1的含乙醇和乙酸的富硫酸盐废水的UASB中,以乙醇为底物(通过SRB)还原硫酸盐的Desulfovibrio物种并没有明显优于以产甲烷(通过MA)为主的Methanosaeta conilli GP6。Chen等[14]为了探索厌氧反应器中菌群的长期竞争动态,在实验室规模的UASB中,恒定COD/SO42-为1的条件下,连续运行329天,处理含乙醇和乙酸的富硫酸盐废水。并通过设计一个全面的结构化数学模型来扩展厌氧消化模型1 (ADM1),预测和探究硫化物和产甲烷菌对电子的长期动态竞争。在整个过程中明显出现了MA优势阶段、相持阶段和SRB优势阶段。从而说明,SRB在水处理环境中的竞争行为是一个长期且阶段分明的过程,可以通过研究介导这个过程中的环境条件、菌群构成,从而对竞争结果进行干涉和改变,使竞争向有利的方向进行。
在微生物电化学过程中,SRB和MB作为两种重要的微生物群落。它们之间的长期竞争对废水处理效率和稳定性具有重要影响。微生物电化学技术在处理含硫酸盐废水中可以利用微生物和固体电极的相互作用来实现污染物的去除,而在这一过程中,SRB和其他微生物会在电极表面展开竞争,或通过电子传递过程及代谢活动参与硫酸盐的降解;另一方面,通过电化学调控,适当地施加电场,可以促进优势菌群的生长,从而对竞争结果进行改变或加速整体的竞争进程。同时微生物电化学还原硫酸盐的主要产物是硫化物,而硫化物是一种排放要求更严格的污染物。因此,硫酸盐污水的完整处理应整合硫化物的氧化回收过程。Pozoetal团队[15]通过构建富集硫氧化菌的生物阳极对硫酸盐还原出水进行处理,取得了74%的硫回收率。因此,生物电化学不仅能应用于废水中硫酸盐的还原,还能应用于硫酸盐废水中单质硫的回收。
2.3 不同环境下硫酸盐还原菌的竞争对象
表1 不同含硫酸盐废水处理过程实例Table 1 Examples of treatment processes for wastewater with different sulfate content |
| The role of SRB in different water treatment processes | Sulfate concentration in the system (mg/L) |
|---|---|
| Using BES to treat high-strength sulfate wastewater (1000 mg/L) without external electron donors and recover elemental sulfur[16]. | The concentration of sulfate in wastewater is 1000 mg/L. |
| The integrated system composed of anaerobic SRBR and SOFC is applied to treat high concentration sulfate/sulfide (1300 mg/L) organic wastewater. The sulfides produced by SRB during sulfate reduction process not only act as endogenous electron media, but also act as electron donors to oxidize sulfides into non-toxic sulfur, which can be recovered through precipitation[17]. | The concentration of sulfate in wastewater is 1300 mg/L. |
| In the UASB reactor, NH4+oxidation is driven by SO42- reduction, where anaerobic ammonia oxidizing bacteria dominate denitrification, SRB and SOB participate in sulfur cycling, and receive the electrons required by anaerobic ammonia oxidizing bacteria through direct interspecies electron transfer (DIET) pathway[18]. | NH4+/SO42-:0.8 (NH4+-N:70 mg/L, SO42-S: 80 mg/L) |
| During the anaerobic digestion process of slaughterhouse wastewater sludge at medium temperature, the high pH value of the initial sulfur-containing sludge (with a sulfate content of 500 mg/L) inhibits sulfate reducing bacteria, which is beneficial for the growth of methane producing bacteria[19]. | The concentration of sulfate solution in sulfur-containing sludge digestion is 500 mg/L. |
| During the 330 day operation of UASB in treating sulfate rich wastewater, methane production gradually decreased and sulfate reduction efficiency continued to improve. Only after complete reduction of sulfate can MPA convert the remaining COD into methane[20]. | The sulfate content in wastewater is 3000 mg/L. |
| The two-phase anaerobic process was used to start at low load for 33 days. At COD/SO42-~3 (sulfate content of mg/L), the sulfate removal rate in the effluent decreased with the decrease of HRT[10]. | The first stage operation dosage is 500 mg/L, and the second stage is 1000 mg/L. |
| Anaerobic co digestion was carried out on kitchen waste (FW) and waste activated sludge (WAS). When the sulfate concentration is 50, 100, and 400 mg S/L, the co digested methane production is not affected, but significantly decreases at 200 and 300 mg S/L. The presence of sulfates can stimulate sulfate reducing bacteria to convert propionic acid into acetic acid, providing another metabolic pathway for methane production[9]. | The best processing efficiency is achieved when the concentration is greater than 50 mg/L but less than 200 mg/L. 50 mg/L ≤ Sulfate concentration ≤ 200 mg/L |
表2 硫酸盐还原、产甲烷和产乙酸反应Table 2 Sulfate reduction, methane production, and acetic acid production reactions |
| Equation | ∆Gº (kJ/reaction) |
|---|---|
| Methanogenic reactions | |
| 4H2+HCO3-+H+→CH4+3H2O | -135.6 |
| Acetate-+H2O→CH4+HCO3- | -31 |
| Acetogenic reactions | |
| Propionate-+3H2O →Acetate-+HCO3-+H++3H2 | +76.1 |
| Butyrate-+2H2O→2Acetate-+H++2H2 | +48.3 |
| Lactate-+2H2O →Acetate-+HCO3-+H++2H2 | -4.2 |
| Sulphate-reducing reactions | |
| 4H2+SO42-+H+→HS-+4H2O | -151.9 |
| Acetate-+SO42-→2HCO3-+HS- | -47.6 |
| Propionate-+0.75SO42- →Acetate-+HCO3-+0.75HS-+0.25H+ | -37.7 |
| Butyrate-+0.5SO42- →2Acetate-+0.5HS-+0.5H+ | -27.8 |
| Lactate-+0.7SO42- →Acetate-+HCO3-+0.5HS- | -80.2 |
SRB可分为产丙酮类SRB (ASRB)和产甲烷类SRB (MSRB),当环境中的硫酸盐浓度不足以支撑SRB将有机物完全氧化时,在生态学上,SRB种群之间将会彼此进行硫酸盐的竞争,但目前对其种群间的竞争进行解释和探究的研究较少。在硫酸盐限制下,硫酸盐还原菌优先使用H2、乳酸和乙醇作为底物,而不使用丙酸盐和乙酸盐。
虽然SRB是以硫酸盐作为终端电子受体的能力来命名的,但还可以使用其他电子受体进行生长,并且可以在没有有机电子受体的情况下发酵底物。因此,在一个环境中出现大量SRB并不一定反映该环境中发生硫酸盐还原。所以从热力学上判断,硫酸盐还原反应过程从侧面反映着环境中菌群的生存关系,但菌群关系调控是一项复杂且存在诸多不可控因素的工作,此时通过采用额外的干扰,人为影响热力学进程,将能为这项工作提供新思路。微生物电解池(microbial electrolysis cell,MEC)则是一类通过外加电势来实现反应发生的能量净输入系统。因为在MEC中,阴极表面还原电势低于阳极表面氧化电势,系统电势差小于0,系统吉布斯自由能(∆Gº)大于0,故从热力学角度系统反应不会自发发生,需要外加一定的电势,弥补系统存在的电势差来引发电极表面氧化还原反应的发生。阴极硫酸盐还原的电子传递机制主要分为两种,一种是SRB利用在阴极产生的H2还原硫酸盐。Liu等[22]利用碳布作为阴极电极材料,发现当阴极电势为-0.8 V (vs NHE)条件下可产生氢气还原硫酸盐。另一种是SRB直接接受阴极的电子还原硫酸盐。Yan等[23]利用Desulfopila corrodens IS4在纯菌体系下以石墨掺锗电极为电子供体,当阴极电势低于-0.4 V (vs NHE)时,Desulfopila corrodens IS4能够直接接受阴极的电子还原硫酸盐。在混菌体系下SRB能够直接接受电子还原硫酸盐。基于此,MEC也可应用于加速一些热力学上能自发进行但反应速率很慢的反应[15],通过不同功能生物阴极的构建,在电化学驱动下,N、S、C元素的相互关系和与此相关的菌群关系将得到干预和调节。
由于SRB存在的不同菌群关系复杂且多样,在一个体系中对菌群间的相互作用进行完全调控当前是无法实现的。更多的是通过对不同体系,在不同环境影响下对现存的菌群作用进行整理、归纳和分析,并作为类似处理工艺的参考,生物电化学的利用则为调节菌群生长进程提供了新的方式和手段,使研究能有更多的发展可能。
3 硫酸盐还原菌的电子供体利用途径
3.1 热力学上SRB的电子供体利用
硫酸盐还原微生物(sulfate-reducing microorganisms ,SRM)是厌氧原核生物,主要利用异化硫酸盐还原来储存能量和代谢生长。由于硫酸盐接受电子后形成硫化物所产生的能量要比其他途径少得多。尽管硫还原途径具有能量劣势,但SRM已在各种环境中被检测到,并对硫和碳循环产生重大影响[24]。如图4所示,不同种类的SRB构成了自然界中的硫循环过程。从而说明,在水处理过程中,研究人员往往需要对体系中的硫酸盐进行还原去除,但同时也希望控制硫化物的排放,这些过程都离不开SRB,所以对于菌群行为的研究是有必要的。根据地球微生物学中的“热力学阶梯”概念,SRM难以和具有更高能量输出的微生物(如反硝化菌或铁还原菌)竞争相同基质进行生长代谢,从表3可以看出,从热力学上讲,微生物在NO3−存在下无法有效地还原SO42−。但在水处理过程中,脱硫杆菌不仅可以利用硫化物进行氧化还原反应,通过其转化为硫酸盐来去除水体中的污染物质;还可以将硝酸盐还原为氮气,从而减少氮的含量。
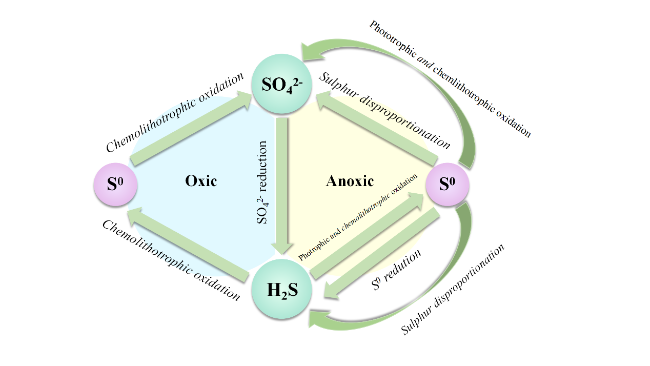
表3 硫酸盐和硝酸盐在不同电子供体下的反应[30⇓~32]Table 3 Reaction of sulfates and nitrates with different electron donors[30⇓~32] |
| Reactions | ∆Gº ((kJ/reaction) |
|---|---|
| Sulfidogenic reactions | |
| 0.125CH3COO-+0.187H++0.125SO42- →0.125CO2+0.125HCO3-+0.0625H2S+0.0625HS- +0.125H2O | -6.18 |
| 0.125SO42-+0.5H2+0.187H+ →0.0625H2S+0.0625HS-+0.5H2O | -19.4 |
| 0.0625C6H12O6+0.126SO42-+0.187H+ →0.25CO2+0.0625H2S+0.0625HS-+0.25H2O | -20.84 |
| 0.083CH3CH(OH)COO-+0.125SO42-+0.187H+ →0.166CO2+0.0625H2S+0.0625HS-+0.25H2O | -11.83 |
| 0.25SO42-+H2+0.25H+→0.25HS-+0.5H2O | -10.68 |
| CH4+SO42-→HCO3-+HS-+H2O | -49.49 |
| Nitrate-reduction reations | -2720 |
| NO3-+3H2+0.23CO2+H+ →0.48N2+0.046C5H7O2N+3.4H2O SO42-+4.2H2+0.015NO3-+0.075CO2+1.515H+ →0.5H2S+0.5HS-+4.17H2O+0.015C5H7O2N 0.0416C6H12O6+0.2NO3-+0.2H+ →0.25CO2+0.35H2O+0.1N2 |
许多行业,如印染、制革和制药等会产生大量同时含有氨(NH4+)和硫酸盐(SO42-)的废水,为对废水中的氨氮进行去除人们开发了氢自养反硝化和厌氧氨氧化等工艺。反硝化菌在氢自养反硝化过程中使用H2作为电子供体,当有SO42-共存时,自养硫酸盐还原将可能成为氢自养反硝化的竞争反应。对于同时含有NH4+和SO42-的工业废水,硫化物自养反硝化是一个不错的解决方案,该工艺使用S2-作为电子供体来减少NO3-将硫化物自养反硝化工艺与异养硫酸盐还原相结合,用于处理含SO42-和NH4+的城市污水[25,26]。采用SO42-—S2-— SO42-循环作为电子传递途径促进反硝化,NO3-去除率可达90%以上,但该方法通常会受到碳氮比不足的限制。而Ren等[25]在SANI工艺中将生物电化学硫酸盐还原与异养硫酸盐还原和硫化物自养反硝化相结合,以去除含NH4+和SO42-的工业废水中的氮。在这个耦合系统中,电子通过硫循环提供给反硝化过程。实质上采用多腔室反应器或使用多个反应器进行串联处理,从而为不同的菌种创造优势条件,避免菌群的竞争,实现协同作用处理污废水[27]。
硫酸盐存在时, SRM在缺氧环境中对H2和乙酸盐有较高的亲和力,这分别是氢营养产甲烷菌和同质产氢菌的主要底物,与MPB相比,SRM对H2和乙酸具有更高的亲合力和更低的阈值[28,29]。所以在水处理中更多的是通过调节SRB与MPB之间对于底物的竞争,使反应器达到稳定平衡,从而获得最为高效的处理效率和能量回收。从而,热力学只能作为对SRB菌群竞争的简单分析,在热力学上虽然反硝化菌相较于SRM更具有优势,但在实际的应用中,硫酸盐还原工艺和反硝化工艺可以通过改进电子传输途径而达到相辅相成;而另一方面,SRB在热力学上虽优于产甲烷菌,但这并不意味着在SRB优势的环境中会对产甲烷菌产生全面的抑制。相反,SRB可以通过对长链脂肪酸的利用间接促进产甲烷行为。
生物电化学的引入,则为SRB的菌群竞争提供了新的思路。基于MEC与传统的厌氧反应器耦合的生物电催化反应器处理高浓度硫酸盐废水,通过阳极氧化有机物,实现有机酸的快速分解,从而维持系统内的酸平衡,有利于提高厌氧消化效率,提高整体反应器的抗冲击能力。此外,阴极作为电子供体可强化硫酸盐的降解能力。因此,将MEC降解污染物和厌氧硫酸盐还原降解污染物相结合,理论上具有更高的污染物降解能力,有望加快降解硫酸盐废水中的有机物。
3.2 电介导对SRB菌群的影响
根据对碳源的利用,SRB会被异化或同化,前者是异养型SRB,以有机化合物为底物,后者是自养型SRB,以CO2为碳源,以H2为电子供体。含硫工业废水处理多利用异养型SRB,这类SRB不仅与MB具有竞争作用,而且与产乙酸菌具有协同作用,但是否与自养型SRB对CO2竞争还未见明确报道。自养型SRB可以在H2、CO2和CO作为唯一的电子供体的情况下生长[33]。Houten等[34]以H2和CO2为底物,发现中温生物反应器在10天内就能达到较高的硫酸盐还原率。 研究表明,以合成气(H2、CO2和CO的混合物,以及少量甲烷和氮等其他组分)为电子供体,7周后硫酸盐转化率可以达到15 kg/(m3· d),硫酸盐的平均去除率为88% [35]。采用合成气作为电子供体不仅具有经济效益,而且处理出水不会存在有机物残留。然而,现今对合成气的利用受限于H2转移速率,同时还需要考虑SRB与其他微生物的竞争和由于甲烷生成所导致的H2利用效率低等问题[36]。在处理含氨氮废水中,异养和自养细菌之间的协同作用被用于提高反硝化效率,其中异养代谢产生的CO2可以作为自养细菌的碳源。然而,迄今为止,尚未观察到异养型SRB和自养型SRB之间存在明显的协同作用。
无论是基于对不同碳源的利用,还是以对底物的氧化程度进行分类分析,都与SRB对电子供体的利用和不同的电子传递途径有关。在自然条件下,菌群的竞争结果受到环境和反应热力学的影响,但生物电化学引入后,通过外加电场的作用来驱动或加速一类化学反应成为可能。此外,如图5所示可通过在体系中添加导电介质,促进种间电子转移,从而改变菌群的代谢,为实际的污废水处理提出了更多的可能性。基于多种微生物物种之间的能量和电子传递的共生关系,其中化能异养细菌和MA之间的共生作用对在不同厌氧环境下生产CH4有很大贡献,同时对全球碳循环也有很大的影响[38,39]。例如,在电合成产甲烷反应中,从化能异养细菌到产甲烷古菌的种间电子转移不仅能通过作为电子载体的小分子(例如H2和甲酸酯)扩散发生[40,41],而且可以由通过导电固体材料的电流介导,这一过程被称为电同步或直接种间电子转移[ 42,43]。
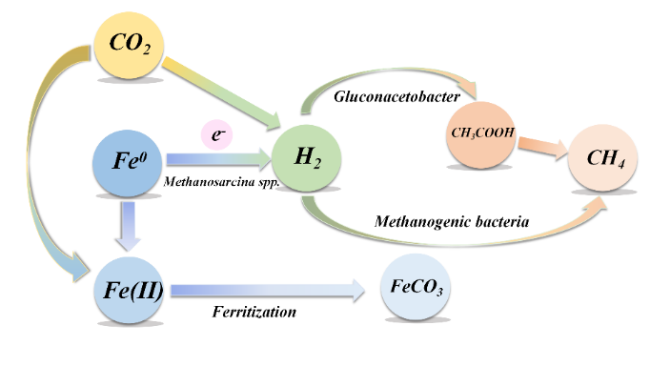
导电介质的添加主要影响了环境中的电化学作用、聚集作用和微生物活性。在产甲烷系统中,为了促进甲烷的生产,提出了补充导电颗粒作为改进厌氧废水处理系统的新型生物技术[44⇓~46]。在最近的研究中,将零价铁(ZVI)添加到AD系统中可以促进CO2的原位生成CH4,从而通过生物技术实现碳捕获,在体系中同时补充水合铁和硫酸盐时促进了甲烷的生成,硫酸盐和氧化铁的共存可以通过(半)导电硫化铁的生物矿化促进甲烷的生成,通过电合成作用产生甲烷。从菌群角度看,在同时添加水合铁和硫酸盐时,富集的DIRB和SRB菌群改变了系统中的代谢途径和电子流,从而促成了MB的富集并提高了CH4的生成速率[47]。在金属废水处理过程中,常采用添加氧化石墨烯(GO)、还原氧化石墨烯(rGO)、ZVI等,图6所示,ZVI和SRB协同作用去除废水中的重金属。导电颗粒在水中所具有的良好导电性使其能参与到不同的电化学反应中。在电解质溶液中,导电颗粒表面会发生氧化还原反应,并产生电流,这种电化学作用促进了水中污染物的氧化、还原或沉淀,提高了废水处理效率。
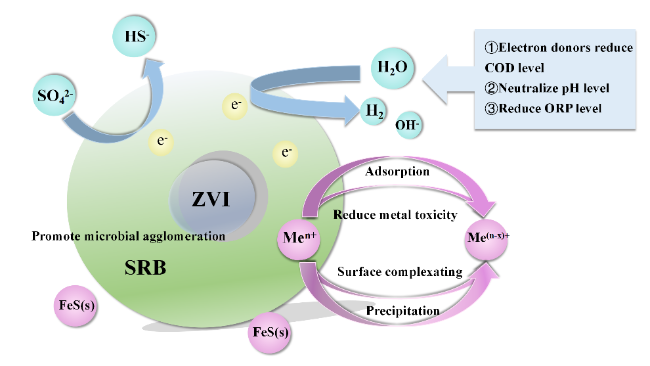
ZVI的加入可以显著提高SRB的活性和SRB系统中重金属的去除率。ZVI作为电子供体,减少了MB和SRB在含硫酸盐废水中对电子的竞争,附着在ZVI上的SRB可以从ZVI腐蚀过程中产生的H2中获得电子。这降低了硫酸盐还原的COD[48⇓⇓~51]。ZVI-SRB对重金属的去除主要依靠还原沉淀、硫化物沉淀、共沉淀和生物吸附[52]。此外,ZVI的加入改变了富硫酸盐环境中的微生物成分,对水中菌群竞争起到了缓冲剂的作用,同时也为微生物提供了良好的附着表面,促进生物膜的形成。导电介质的出现使得对菌群关系调节的影响条件不仅局限于环境因素,如表4所示,不同的导电介质适用于不同的水处理环境,而电化学的引入,不仅影响了体系内菌群的构成,同时也改变了细菌的电子传输途径,在面对更复杂多变的水环境时,提供了更丰富灵活且能量友好的处理方法。
| Non biological conductive materials | characteristic |
|---|---|
| Iron oxide | Different types of iron oxide particles have different conductivity. The conductivity of iron oxide can determine the methane production rate of microbial communities. (Semi) conductive iron oxide can promote methane production of acetate and ethanol, and improve lag time and yield. |
| Biocarbon and activated carbon | Biochar provide substrate for microbial adhesion and attachment, and reduce inhibition by absorbing possible inhibitors in Anaerobic digestion. Biochar mediated DIET promoted the degradation of organic acids. Different particle sizes of GAC and powdered activated carbon (PAC) have an impact on the enhancement of methane generation |
| Carbon fibre | The carbon cloth and felt have good conductivity, and the biofilm is easy to form on the surface of the carbon fiber, which helps to increase the residence time of biomass in the reactor, thus improving the AD performance. The carbon cloth can alleviate the acid shock and high hydrogen partial pressure in the Anaerobic digestion. |
| Graphene | It has unique physical and chemical properties, such as extremely high conductivity, large surface area, and high mechanical strength. After adding Graphene, the activity of the main coenzyme F420 in the process of methane production was significantly increased. A smaller amount of Graphene is required for DIET stimulation. |
| Carbon nanotubes | Carbon nanotubes can promote symbiotic co cultivation and methane generation in complex environmental matrices, but their economic costs are high. |
3.3 SRB节能产氢途径
SRB在厌氧环境中是主要的耗氢微生物,但在缺乏硫酸盐的环境中,SRB可以作为产氢微生物在体系中发挥作用,这为开发基于SRB的相关处理技术提供了广泛的可能。值得指出的是,SRB具有极高的氢化酶活性并同时具有利用乳酸、乙醇、甲酸盐和丁酸盐的能力[54]。这意味着在硫酸盐有限的环境中,可以通过与其他生物合胞产生H2、CO2和醋酸盐进行发酵生长[55]。Odom和Peck[56]提出的硫酸盐还原过程中可能产生H2的中间产物也表明,在没有硫酸盐的情况下,SRB可以作为H2生产者。生物氢(BioH2)生产是一种非常有趣的能量输出,其只需要较低的能量投入,并且如果使用废物或可再生物质作为底物,那么这个过程将是可持续的[57,58]。这展现了SRB在节能产氢上有着不容忽视的潜力。
氢化酶是SRB中一种重要的电活性酶,通过催化氢的析出和氧化能将其活性与固体电极结合起来[59]。研究主要集中在利用这些细菌作为MEC中的生物阴极,SRB的电催化制氢活性大于或等于它们的H2氧化活性,这证实了它具有较高的制氢潜力[60]。MECs近期成为一种新颖的、有前途的可再生制氢技术,通过在环境压力和温度下施加外部电压的微生物电源,MECs能够提供将H+还原为H2所需的能量[61]。Desulfovibrio sp.是MEC生物阴极中的优势菌属,利用该菌属提出了一种利用阴极上的电子来减少质子的氢循环机制。存在于细胞质膜或细胞质中的氢化酶通过酶促反应来减少质子。而一旦细胞质质子被消耗,氢则会通过细胞质膜扩散到外周质,此时氢将被氧化,质子驱动力就会启动。质子驱动力则可以通过流向细胞质的电子来维持,这减少了质子对硫酸盐的消耗[62]。研究发现以甲基紫素为介质的纯培养脱硫弧菌可以产生氢气[63]。有研究表明,Desulfovibrio sp.可以通过参与存在于膜细胞质上的氢化酶或节能氢化酶来保存能量[64]。在SRB产氢系统中,因为SRB作为产氢菌时的效率优于MA(厌氧发酵过程中的微生物聚集体,在产氢反应中起着重要的作用),所以通常使用BES抑制产甲烷菌以及从厌氧发酵残留物中回收氢气。客观来说,MB是几乎无法利用丙酸作为代谢底物的,当MEC中产生丙酸减少的现象时则说明此时存在微生物电解的情况[65],但较高的丙酸/乙酸比将会导致厌氧消化过程失败,从而可以通过抑制丙酸型耗氢途径提高产氢量,这也可能有利于SRB混合培养的氢气回收[66]。
其中,铁作为一种重要的营养物质和一种特殊的电介导材料,不仅是刺激氢营养型产甲烷菌生长所必需的,也有助于促进其他金属的利用[67]。在UASB反应器中,添加10 µmol/L Fe可使污泥的产甲烷活性增加一倍[68]。通常,Fe和Ni以Ni-Fe-s簇和Fe-Fe-s簇的形式存在,形成酶亚基,包括氢化酶和乙酰辅酶a合成酶[69]。Fe的添加还影响了硫酸盐还原菌的代谢途径从而对SRB产氢过程产生促进作用,Fe可以作为一种电子传递体参与菌体内的电子传递过程,从而促进产氢反应的进行。此外,Fe还可以提供菌体所需的酵素和辅酶的构建材料,进一步增强菌体的产氢能力。如前文所提,通过添加适量的铁元素,可以优化系统中菌群的生长环境,缓解电子供体的竞争,从而促进系统中的处理平衡。
在自然环境中,多种电子供体都能为SRB所利用,在传统手段中,可以根据热力学优势,判断体系内SRB对不同底物的氧化程度从而人为地进行关于SRB的菌群筛选。而在本质上,则是对SRB电子供体利用程度的研究。区别于单纯通过营养底物进行菌群关系调控,通过投加电子介质或通过使用电极作为电子供体来达到改变电子传输途径的方法,无论是在水处理工艺还是在产氢节能上,都能获得不错的效果。
4 硫酸盐还原菌的调节控制因素
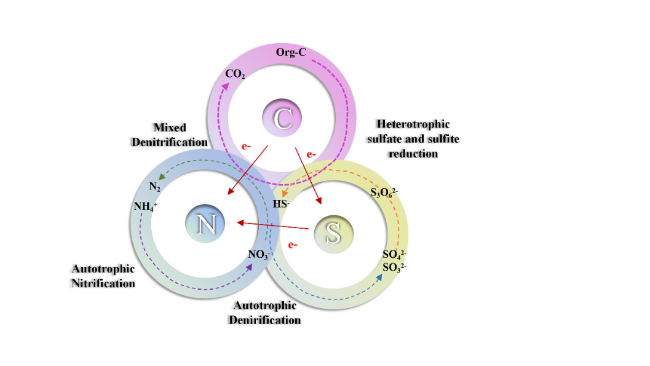
细菌的代谢途径和SRB对于电子供体的利用途径对单个反应器中生物多样性和菌群关系方面起着重要作用,而SRB的多样性直接或间接地也受到了外加电场、导电介质、碳源类型及碳硫比、水力停留时间(HRT)、pH值和温度等工艺参数的影响。同时,硫作为一种常见的自然元素,与其他元素循环存在着密切关系,如图7所示,而其中硫酸盐还原微生物在对氮、磷的无害化去除方面也发挥着重要作用。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
4.1 外加电场的影响
外加电场的引入,实质是引入了微生物电化学反应过程。电压的施加主要是通过增强甲烷生成和硫化物生成,以有效处理富含硫酸盐的废水。适当地施加电流会增强细胞外分泌,不仅有利于生物膜的形成,也增强了向MA的电子转移。在微生物电化学系统中SRB可以触发强有力的协同作用,微生物分析表明,电场的存在明显增加了系统中脱硫弧菌(Desulfovibrio)的丰度,Desulfovibrio可以通过菌毛向甲烷古菌和甲烷细菌直接传递电子。此外,适当的电刺激有助于促进功能性酶和微生物代谢,研究通过对施加电场(EV)反应器和未施加电场的反应器进行硫酸盐去除对比实验,同时对两个反应器内的COD/SO42-比值(CSR)逐渐降低,与对照组相比,EV反应器在CSR为2.0时,甲烷产量增加了30%,硫酸盐去除率增加了40%[70]。其次,微生物电化学系统可以维持SRB系统的ORP(oxidation-reduction potential)环境,并且电场的存在可以减少底物的消耗。从理论上解释,当细胞膜带电时,内膜电性为正,外膜电性为负[71]。电场对微生物的影响可能主要在两个微生物生理过程上:(1)电压敏感通道的激活;(2)更高振幅的电穿孔对细胞膜的渗透[72]。但有研究指出施加的电流会影响Na+的浓度,随之膜可能因为电压作用打开Na+通道而去极化,这将可能对细胞产生负面影响[73]。因此,微生物电化学系统中的微生物将会产生两种不同的变化。一方面,电流刺激细菌的活性和污染物的降解。另一方面,随着膜通透性的改变,特别是物质和能量通道的无序化,过高的电流会抑制微生物的活性,甚至导致细胞膜破裂。有研究表明在富硫酸盐MEC中最佳应用电流为1.5 mA,施加电压使硫酸盐去除效率提高14.9%。但过大的电流会导致细胞膜破裂,导致生长和代谢活动减慢,从而导致硫酸盐去除效率降低,而适当的施加电流可以促进细胞外分泌的产生,有利于细菌在阴极生物膜中的富集;阴极上生物膜的形成可以加速阴极的氧化[74]。
电场的施加同时影响着氧化还原电位的改变,过高的电压施加产生的电解作用甚至会改变体系的环境,从而对体系中的微生物群落造成严重影响。此外,在厌氧水处理过程中,一些氧化还原物质会对ORP产生影响,如NO2-、NO3-和ZVI等[78]。这也证实了导电介质的加入不仅影响了电子传递途径,而且其自身的理化性质也会对体系造成影响。
4.2 导电介质
导电颗粒在废水处理中的应用主要包括电化学方法和生物处理方法。在电化学方法中,导电颗粒可以作为电极材料参与废水处理过程;而在生物处理方法中,导电颗粒则可以作为载体促进微生物的附着和代谢。在生物电化学方法中,导电颗粒如纳米碳粉、铁粉等通常被用作电极材料,参与废水处理过程。添加导电颗粒可以提高电极的导电性能和活性表面积,增强电化学反应的效率。例如,添加碳纳米管到电极表面可以显著提高电极的电化学活性,并促进废水中有机物的降解[79]。在生物处理方法中,导电颗粒能够为载体提供良好的附着表面,促进微生物的附着和代谢活性,从而增强废水中有机物的生物降解效率。将碳纳米管等导电颗粒引入生物颗粒污泥中,可以显著提高微生物对废水中有机物的降解速率和稳定性[80]。在UASB反应器中添加颗粒活性炭(granular activated carbon,GAC)增加了高浓度硫酸盐有机废水的处理效率,与未添加介质相比,GAC增强了微生物活性,改善了潜在的电活性微生物与MB之间的直接种间电子转移,同时也提高了微生物的代谢活性,包括糖酵解、丙酮酸代谢、硫酸盐还原和甲烷生成。因此,关键微生物的富集、促进DIET产甲烷、提高功能代谢活性可能是GAC反应器污泥产甲烷活性较高的重要原因[81]。导电颗粒的添加可以通过增强电化学反应的效率或促进微生物活性,从而提高废水处理的效果。颗粒活性炭(GAC)或磁铁矿可以促进有机废物产生甲烷,其中磁铁富集了污泥中的铁还原菌,而颗粒炭具有较高的电导率和较大的比表面积,促进了铁还原菌与MB之间的共营养代谢。在厌氧消化池中同时投加磁铁矿和GAC,可同时加速污泥的水解和甲烷的产生,从而获得更好的污泥消化性能[82]。
在实际运用中,需要根据废水特性和处理工艺选择合适的导电颗粒及其添加方式,同时导电颗粒的添加还可能会对废水处理系统所需的电流密度、电位范围和pH值等参数产生影响。在BES中,通过调节氧化还原电位可以影响导电颗粒参与废水处理过程的效率。例如,在MFC中,调节阳极和阴极之间的氧化还原电位可以影响导电颗粒的电子转移速率和电化学活性。在双腔MEC中添加磁铁矿颗粒从而对体系中的自养硫酸盐还原生物阴极起到了增强作用,使硫酸盐还原率提高了85%,并且阴极上的生物量、生物膜厚度和SRB菌群丰度与未添加磁铁矿颗粒的反应器相比有了很大的提高[83]。通过调节阳极和阴极之间的氧化还原电位,还可以优化导电颗粒的电子传递效率,从而提高废水处理的效率[84]。pH值对于废水处理中微生物代谢活性和电化学反应的进行都具有重要影响。在添加导电颗粒的生物处理过程中,调节废水的pH值可以影响微生物与导电颗粒的协同作用,进而影响有机物的降解效率。研究发现,在不同pH条件下,导电颗粒对废水中有机物的降解速率和降解途径可能存在差异[85]。温度是影响微生物代谢活性和电化学反应速率的重要因素。在添加导电颗粒的废水处理过程中,调节温度可以影响微生物的生长速率、代谢活性以及电化学反应的速率。在不同温度条件下,导电颗粒对废水中有机物的降解效率和循环利用效果会有所不同[86]。这些操作条件应根据具体情况进行调整,以实现最佳的处理效果,并结合实际情况进行调整和优化。
4.3 碳源(类型及碳硫比)对SRB 的影响
Lens等[83]报道了SRB在碳源利用和代谢活性方面的多样性。不同碳源转化途径的差异是影响硫酸盐还原过程的主要因素,这直接影响了完全氧化SRB (CO-SRB)和不完全氧化SRB(IO-SRB)的竞争(还影响了反应器的启动时间,因为IO-SRB更容易富集,也同时影响了SRB的还有碳源的量,但在实际应用中,SRB需要和环境中的微生物进行碳源竞争)。由于IO-SRB的存在,糖和醇将快速不完全氧化产生乙酸,导致溶液pH降低抑制SRB的活性;相反,通过转化pH稳定的小有机酸则会产生更多的硫化物[88]。因此,当使用不同碳源作为基质底物时,体系中占优势的菌群类型也会随之改变,例如,以NaHCO3和乙酸盐为碳源的SRM以Desulobacter和Desulfobulbus为主,以乙醇为碳源的硫酸盐还原菌以Desulfoicroum和Desulfovibrio为主[89]。而SRM对于不同底物的转化速率也是存在差异的,底物消耗速率取决于电子供体和硫酸盐的浓度,这也必然会影响SRB和MB之间的竞争。如表5所示,SRB在不同碳源利用存在着热力学的差异。不同碳源可以在两个方面对微生物硫酸盐还原产生影响:一方面,SRB对不同的碳源具有不同的亲和力;另一方面,SRB的碳源代谢过程可能通过影响溶液的pH和ORP对SRB活性产生反馈。由于实际水环境中会存在着多种基质,从热力学上,不同菌群能使利用最优化的能量源也是不一样的,能量利用最大化也会使体系中的竞争行为最小化。在实际应用中,可以根据特殊污染物类型或需要的产能类型来决定水处理体系中的优势菌群类型,并通过碳源的投加类型和浓度进行调节。
表5 硫酸盐还原反应中不同电子供体的热力学[90]Table 5 Thermodynamics of different electron donors in sulfate reduction reaction[90] |
| Electron donors | Product | ∆Gº (kJ/reaction) |
|---|---|---|
| 4H2+SO42-+H+ | HS-+4H2O | -151.9 |
| 4Formate-+SO42-+H+ | HS-+4HCO3- | -146.7 |
| Acetate-+SO42- | HS-+2HCO3- | -47.3 |
| Propionate- +SO42-+H2 | HS-+HCO3-+Acetate-+H2O | -75.8 |
| Propinate- +2SO42-+H2 | 2HS-+3HCO3-+H2O | -122.7 |
| Butyrate- +3SO42-+2H2 | 3HS-+4HCO3-+5H2O | -198.4 |
| Butyrate- +SO42-+2H2+6H2O | HS-+2Acetate- | -103.8 |
| 4Methanol+3SO42- | 3HS-+4HCO3-+4H2O+H+ | -361.7 |
| 2Methanol+SO42- | HS-+2Formate-+H++2H2O | -108.3 |
| 2Ethanol+SO42- | 2Acetate-+ HS-+2H2O+H+ | -132.7 |
| Glucose+SO42- | HS-+2Acetate-+2HCO3-+3H+ | -358.2 |
| Glucose+3SO42- | 3HS-+HCO3-+3H+ | -452.5 |
| 2Lactate-+SO42- | HS-+2Acetate-+2HCO3-+H+ | -159.6 |
值得强调的是,在含硫酸盐的水环境中,COD/ SO42-是一项重要的指标,其决定了硫酸盐还原和甲烷生成过程中的电子流并直接影响SRB在微生物群落中的比例。理论上,当COD/SO42-比低于0.67时,SRB可以通过利用有机质将硫酸盐完全降解,在这种情况下,所有的电子都会流向SO42-;当比值大于0.67时,SRB与其他微生物对电子供体的竞争将会变得激烈[91]。即比值在1.7~2.7之间时,SRB和MA之间的竞争非常激烈。实际上,当COD/SO42-为0.5时,SRB还原利用电子总量占总电子流的50%以上[92,93];而当COD/SO42-的比值为2.0及更高时,MA使用了总电子流的80%以上。然而,O’Reilly和Colleran[89]研究发现,在进水COD/SO42-比值为2~16的情况下,SRB物种无法与MA物种竞争醋酸盐。这些结果的差异可能与碳源组成、硫酸盐负荷以及pH、温度、ORP等环境因素的差异有关。
不同的外加碳源可以通过对环境因素产生影响从而改变体系中的优势菌群,而不同的碳源投加量通过改变碳硫比而对电子流走向产生影响。但在大部分实际含硫酸盐污废水中存在着低碳源的现象,一般需通过额外投加有机碳源电子供体来弥补,但随之而来的是处理成本的增加,而在最近的水处理中则主要采用了利用生物电化学技术对含硫酸盐废水进行降解,其中采用MFC和导电介质(如氧化铁、ZVI等)在低碳或无碳条件下进行高效处理的技术得到了开发运用;另外,利用纳米材料或使用光催化技术对含硫废水处理同样受到了大量的关注。
4.4 OLR和HRT
有机负荷率(organic load rate,OLR)在有机废水中表示微生物和有机污染物之间的平衡,必须进行控制,以维持产甲烷和酸化之间的平衡[94]。OLR失衡通常会使水解、产酸、产丙酮和产甲烷等步骤失衡[95]。OLR的增加可以通过缩短HRT来进行缓解,然而,在降低HRT和增加OLR的同时,微生物群落将会发生变化,生长缓慢的产甲烷菌可能会被淘汰[96⇓~98]。此外,当底物充足时,挥发性脂肪酸(VFAs)中间产物的形成率会更高,中间产物的额外积累会显著干扰微生物群落,降低厌氧消化的性能,从而对产甲烷菌的活性产生负面影响[99]。HRT还会影响SRB与其他微生物的关系。在较长的HRT条件下,H2产量增加,这加剧了SRB和MA之间的竞争。例如,较短的HRT降低了水解酸化细菌降解大分子有机物的效率,从而导致SRB利用的降解产物减少。在酸性矿山废水处理过程中,水力停留时间对处理效率有着重要影响,较短的HRT会造成反应器内酸的累计并且导致水中重金属无法及时析出分离;而较长的HRT会因为微生物的大量增值从而耗尽营养元素,同时矿物质过度析出,最终导致处理效率降低[100]。合适的HRT需要根据反应器类型和进水特性来确定。例如在填充床生物反应器中,中性pH条件下的最佳HRT为6 h,而在酸性条件下的最佳HRT增加到20 h。在上流式厌氧污泥床(UASB)和下流式流化床反应器(DFB)中,低HRT有利于提高硫酸盐还原率[101]。
4.5 pH值
根据微生物生长所需的pH值,SRB可分为中性粒细胞和嗜酸性粒细胞。pH值为7.0~7.8是SRB中性粒细胞的最佳还原范围,pH值为7.0~7.5时硫酸盐还原率最大[102,103]。在碱性条件下,水解酸化菌可以产生更多的短链脂肪酸,这些短链脂肪酸被SRB有效利用[104],此时,乙酰营养型SRB优于乙酰营养型产甲烷菌[105]。pH值不仅可以影响非游离硫化物占总硫化物的比例,还可以通过影响SRB的活性来影响硫化物与硫酸盐之间的转化[106]。研究发现 [107],硫化物的化学平衡依赖于pH值。pH值为8时硫化物以HS−形式存在,pH值为6时硫化物以H2S形式存在。在pH值为7.0~7.5时,MB和SRB的生长速度相似;较高的pH有利于MB生长,较低的pH值有利于SRB生长。硫化物对厌氧菌的毒性作用在不同的消化条件下有很大的差异,尤其是pH 值[106]。在适当范围内提高pH值可能是缓解硫对MB生长抑制的有效途径[108]。
适合优势菌群生长的最佳pH值因底物和消化技术而异[23]。研究发现,厌氧污泥消化过程中污泥初始pH值对H2S产率具有重要影响,当初始pH值从6.5增加到8.0时,沼气中H2S含量降低44.7%,甲烷含量增加48.6%,这是因为初始pH影响了SRB和MB之间的竞争,而较高的初始pH 值抑制了SRB的生长,从而降低了H2S的产量[19]。此外,污泥pH值显著影响硫化物种类的形成。pH值对SRB的影响是多重的:一方面,pH值直接影响了SRB的生理代谢,控制着细胞内稳态;而另一方面,pH值影响了SRB的生存环境,如环境中硫化物存在形式和脂肪酸的类型,这间接改变了环境中微生物的代谢途径,从而对SRB的共生和竞争关系产生影响。
4.6 温度
SRB可以在很宽的温度条件下生存,但大多数目前已知的SRB主要生存在中温带。中等耐温SRB的适宜温度为40~60 ℃[109]。嗜热SRB的最佳生长温度为65~70 ℃,而80 ℃以上的嗜热SRB仅存在于海洋热液中[108]。适度增温对SRB和MA都有利,但在不同的温度条件下,它们的竞争效应是不同的。SRB在高温废水处理中比MA更具优势。Omil及其团队[108]发现当温度升高到55~65 ℃时,硫酸盐的还原比甲烷的形成要快。造成这一结果的原因可能是由于底物的差异,MA和SRB对如甲醇、乙醇、甲酸盐或乙酸盐这些底物的降解能力可能不同。也有可能在温度的影响下,硫化物的形态和浓度的变化对它们产生了不同的影响,这是因为温度会改变H2S在废水中的溶解度,高温会降低H2S在废水中的溶解度,从而降低H2S对环境微生物的抑制作用[109]。
表6 对水环境中SRB的调节因素Table 6 Regulating factors of SRB in water environment |
| Influence factor | Influence | Adjustment method |
|---|---|---|
| External electric field | The application of an electric field not only changes the type of microbial community but also alters the redox potential. | Select the appropriate electric field strength by adjusting the voltage or current magnitude. |
| Conductive medium | It affects the electronic transfer pathway and its physical and chemical properties can also have an impact on the system. It may also have an impact on parameters such as current density, potential range, and pH value required for wastewater treatment systems. | Choosing appropriate conductive particles and adding methods can affect the efficiency of conductive particles in wastewater treatment by adjusting the redox potential. |
| Carbon source | The conversion pathways of different carbon sources affect the reduction process of sulfates, while the metabolic process of carbon sources causes changes in the pH and OPR of the environment. | Different microbial communities have different advantageous carbon sources, which should be selected based on the type of pollutant and the required production capacity. Adjust microbial behavior by adjusting the carbon sulfur ratio. |
| OLR and HRT | It affects the balance between microorganisms and organic pollutants. | Select based on reactor type and inlet characteristics. |
| pH | It affects the metabolic types of substrates and determines the forms of sulfides in the environment. | The optimal pH value suitable for the growth of dominant bacterial communities varies depending on the substrate and digestion technique |
| Temperature | The competitive effect of microbial communities varies at different temperatures. Higher temperatures can also increase the dissolution of hydrogen sulfide in water. | Adjust the temperature according to the type of dominant microbial community. |
在上述各种影响因素中,大多数已知的SRB对弱酸性环境(pH < 5)的敏感性[109]和它们的低生长速度限制了硫酸盐还原系统的设计和应用。虽然可以通过添加石灰或使用侧流SRB反应器来提高pH值以避免SRB与酸性废水直接接触,但低pH值 (pH < 5)的SRB生物反应器的培养和运行仍然缺乏研究和总结。颗粒污泥为缓慢生长的厌氧生物质提供了一种解决方案。自固定化SRB颗粒可以提高SRB系统的效率,因为它可以提高生物质浓度,减少反应器体积,并增加反应器对pH值、温度等波动的耐受。所以,电场耦合作用实则是通过调节包括SRB在内的菌群电子传输途径从而改变竞争结果,为含有硫酸盐和氨氮的废水提供新的处理可能。
5 结论与展望
SRB利用硫酸盐作为终端电子受体降解有机化合物。然而,它们的能量代谢并不局限于硫酸盐还原,SRB可以广泛使用电子受体。在菌群关系上SRB的一个重要生理特征是可以与产甲烷菌竞争或根据硫酸盐的可用性与产甲烷菌共养生长,其中SRB与其他菌群的共代谢灵活性值得进一步关注,这对将SRB用于水处理和产能上有着重要意义。
此外,在缺乏硫酸盐的环境中,由于氢化酶发挥效能,SRB主要作为产氢菌而存在并与其他菌群协同作用产氢。但关于硫酸盐不足条件下SRB产氢效能受菌群竞争影响方面的研究较少,以及对于SRB产氢效能与体系中产甲烷效能的平衡有待研究。所以当SRB作为产氢菌时需要考虑SRB与SMB在电子供体上的竞争行为和竞争调节效益,即在产氢和产甲烷之间存在的效益平衡,同时还需要考虑,SRB的存在对氢营养型产甲烷菌、乙酰营养型产甲烷菌的影响。当前更多的研究围绕于解决SRB菌群的作用下使体系中硫元素的循环来改变电子流走向,从而提高处理效能。但要真正实现在含硫酸盐和高浓度特殊污染物实际废水领域的广泛运用,关于耦合S和C/N循环的实际模型还有待建立。随着微生物电化学的引入,电场的调节和投加不同的导电介质将会对污废水的处理给出不同的处理可能性。而关于外加电场的生物反应器中产甲烷菌群和硫酸盐还原菌的分布、多样性及其相互关系的研究目前尚未有充分的研究。
水环境中的碳、氮、硫元素在细菌对电子供体的利用中不断进行着循环变化,这印证了SRB与其他菌群在体系中的相互作用,借助地球化学的基本理论对菌群竞争进行机理分析,有助于对混合复杂废水的处理提出有效的措施。通常水质特征和环境条件决定了处理体系中的优势菌群,因此相关工艺参数的调整可以对菌群的竞争进行有效的人为调控,并结合分子生物学技术分析预测硫酸盐还原菌在环境中所发挥的效能或所担当的角色。